|
Title: Triassic stem caecilian supports dissorophoid origin of living amphibians Authors: B.T. Kligman, B.M. Gee, A.M. Marsh, S.J. Nesbitt, M.E. Smith, W.G. Parker, & M.R. Stocker Journal: Nature DOI: 10.1038/s41586-022-05646-5 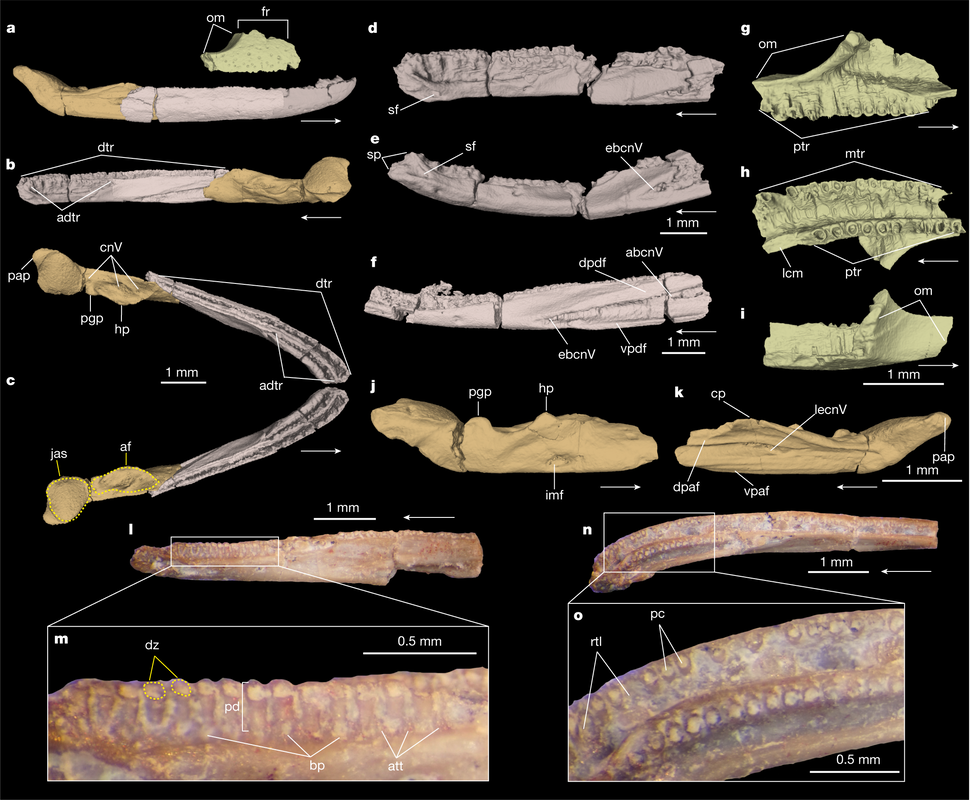 Figure 1 from the paper, showing digital renderings and photographs of the different skeletal elements from Funcusvmeris in different views (go to the article for the full caption, it's way too long to copy in here). General summary: The origin of modern amphibians (frogs/toads, salamanders/newts, caecilians), which are more often termed 'lissamphibians' by scientists to differentiate from the more ambiguous 'amphibians,' has long been a vexing problem. The three modern groups are remarkably morphologically disparate, which makes it hard to both confidently identify and conceive of the ancestral lissamphibian. Lissamphibians today are also relatively small, a pattern thought to have characterized much of their evolutionary history and also an attribute that predisposes their remains to not have been fossilized. Exacerbating this is the extremely poor record of caecilians, which, even today, are a rather cryptic group found only in the equatorial regions and that typically burrow, making them rather hard to observe. Burrowing animals, especially those that not only burrow but that spend much of their lives underground, also have a poor fossil record, which compounds the problems for caecilians. The first caecilian fossils were not even reported until 1972 (previous reports were a misidentifed catfish spine and a misidentified cephalopid, respectively), and to date, there are less than a dozen distinct occurrences of fossil caecilians known globally over an 180 million year interval.
In this study, led by my colleague and current Virginia Tech PhD student Ben Kligman, we report nearly 100 new specimens of the earliest known caecilian in the fossil record from a single Late Triassic site in Petrified Forest National Park, Arizona. Although no complete skulls or skeletons are known, numerous fragments preserve unequivocally diagnostic features found only in caecilians, such as a jaw comprised of largely fused elements that remain separate in other tetrapods and two rows of small teeth with a distinctive feature called pedicelly – a dividing zone at the mid-height of the tooth that often leads the tips to be lost during preservation. This occurrences pre-dates the previous oldest occurrence of caecilians by at least 35 million years and provides new insights into the early stages of the group's evolution. In particular, the new fossils appear to capture the transition toward the modern caecilian condition in which there is extensive co-ossification of multiple elements to form a more consolidated skull (good for burrowing). Their occurrence in Arizona, which was positioned close to the equator in the Late Triassic, suggests that an origin within the equatorial belt also constrained their dispersal, therein offering an explanation as to why caecilians remain tied to these regions when frogs and salamanders have nearly a global distribution except at the poles (and Australia for salamanders). |
About the blogA blog on all things temnospondyl written by someone who spends too much time thinking about them. Covers all aspects of temnospondyl paleobiology and ongoing research (not just mine). Categories
All
Archives
January 2024
|
 RSS Feed
RSS Feed
