|
As many people know, I am an avid sports fan - comes with the territory growing up in L.A., I suppose. Unsurprisingly, I'm also a big fan of sports analysis shows and blogs, which form the inspirational backbone of the last #TemnospondylTuesday post of 2019. One of my favorites is Matthew Berry's Love/Hate column for fantasy football. I essentially grew up without NFL team since both the Rams and the Raiders skipped town after the '94 season (when I was born), and the Rams didn't move back until the 2016 season (when I moved to Canada). As most Canadians know, the CFL is about as popular in Canada as curling is in California, so there's not much going on with that front. So fantasy football is about as exciting as it gets for football; the Rams' current struggles aren't helping either, and does anybody actually know any Chargers fans who live in L.A.? Anyway, Berry often starts off his columns with a story of semi-relevance to his column highlighting which NFL players you should sit or start for the upcoming week, a model that I'll be following in the next section. There is A LOT in this post (perks of spending the post-holiday time cat-sitting for my labmate, Kayla), so buckle up and let's get after it!
I started this blog at the start of the new year, and I have to say, I'm pleasantly surprised that I both managed to keep blogging as semi-regularly as I did and that it got as much traction (a lot not from social media, I might add)! There is clearly an appetite for all things temnospondyl, and I am here for it! Stay tuned as I probably start the year pretty strong on a weekly basis and then fall off the face of the earth for a few weeks in the summer. And keep writing me! Sometimes I don't always respond or I'm real slow to do so, but it's not because I'm getting tired of hearing about how much you like temnospondyls - it's because I have a day job (and a night job) as a PhD student...
When I started out, I wrote up a list of topics that I thought I'd blog about. I've done a few of those, but mostly I've just been spitballing as things started coming to me. I always think you should write what you want, not what you said you would, so here we are. The end of the year auspiciously falls on a #TemnospondylTuesday, thereby imposing an entirely arbitrary sense of particular responsibility to send the blog off on a high note. I thought about doing a summary of temno research in 2019 (meh, sorta predictable) or highlighting stuff I'm working on for next year (too specific). As usual, as I asked my former labmate, Yara Haridy, what she thought I should write. She suggested I do "temnos of the last decade, 2000-2010." After I reminded her that it was in fact about to be 2020, and not 2011, she started spitballing. "What has changed"; "what do we know"; "who's been kicked out of the fam." The alternative prompt was "you just woke up from a coma and you hadn't read a single paper but wanted to be caught up - what do you have to know?" This situation doesn't actually require a coma since most people don't read temno papers. I'm not actually sure most of them want to be caught up either, but they say that overexposure (sometimes) builds an affinity for something, so might as well try it, right? Ten Things I Love, Hate, or Love and Hate
One of my favourite features from the ESPN columns is the one that ESPN senior writer Zach Lowe puts out each week: "Ten Things I Like and Don't Like," which involves a lot of videos (sorry, I don't have any candids of my subject matter), detailed analysis, and give-it-to-me-straight honesty (which can still be complementary). I've come up with my own "Ten Things," which will probably be easy to figure out where each lies on the 'Love-Hate' spectrum and which are listed in absolutely no particular order.
2. No temnospondyl is too good to be cut. So we're not at the level of cutting holotypes just yet, but why bother when your sample size is in the dozens and nobody else cares about 'em? Temnospondyls have been getting cut for a long time; you can go back to Richard Owen who identified the labyrinthine infolding (plicidentine) of Mastodonsaurus that gave the 'labyrinthodonts' their hallmark name. Armand de Ricqlès also cut a lot of different temnospondyl elements back in the day, and now that histological methods and the pre-sampling conservation of target specimens has drastically improved, almost nothing is off limits. Nearly every major clade of temnospondyls has been sampled to date, although it continues to skew heavily toward stereospondyls, which are larger and more often preserved in mass death assemblages. Given that temnospondyls do nearly everything other than take to the skies, workers of other groups looking for a precedent or a comparative data point for more uncommon microanatomical / histological features (e.g., pachyostosis, non-pathological calcified cartilage, thin sections of skulls) should definitely give temnos a thorough look! You can find more on specific studies later in this blog.
3. Not enough independent testing of phylogenetic topologies. I'd like to see more independent workers producing novel phylogenetic matrices to test longstanding (or not so longstanding) topologies. A lot of the existing ones admittedly derive from a small handful of workers and are essentially passed along, nearly unmodified, with simple taxon additions (I'm guilty of this too; it's fast and also nice to assume that previously published work is rigorous). This isn't to say that I think that the base matrices are wrong or bad, but we also won't know how robust they are until somebody else tries to do it from scratch. At least independently rescoring of all sampled taxa to assess the rigor of codings would be nice.
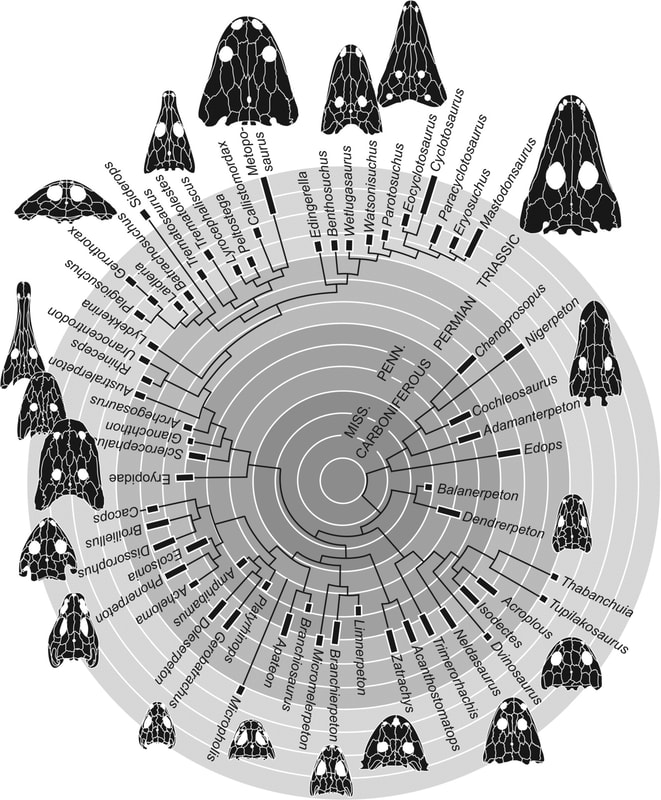
The phylogeny of temnospondyls from Schoch's (2013) analysis mapped onto stratigraphic occurrence; this matrix forms the skeleton for the majority of larger-scale temnospondyl analyses that have followed.
6. Temno paleoart is flourishing!
6B. Browers giving you issues with embedded tweets.
This has nothing to do with temnospondyls, but you'll notice that I embedded a lot of different tweets here. Depending on your browser and ad-block / tracking / cookie settings, you may not be able to view the associated images and instead just see things as a block of text. Sorry...it's not me or my website builder, it's the internet...
7. Participation in temnospondyl research by underrepresented minorities is not flourishing...
No surprises here, but most temnospondyl work has historically been done by white dudes. The earliest workers were guys like Alexey Bystrow, Robert Carroll, E.C. Case, E. D. Cope, Al Romer, D.M.S. Watson, and Samuel Williston, and all of those guys were from Europe or North America. Margaret Steen was one of the few prominent women to work on temnospondyls in the early 20th century, with a particular focus on coal swamp specimens (e.g., Steen, 1930, 1934, 1938). Temnospondyl research hasn't really diversified much in the late 20th century, whose most prominent workers were still dudes, such as Dave Berman, John Bolt, John Cosgriff, Andrew Milner, Ivan Novikov, and Mikhail Shishkin, although Eleanor Daly did her PhD dissertation (1981) on what ended becoming Eoscopus locklardi, an amphibamiform from Kansas, and both Anne Warren (now emeritus at LaTrobe University in Australia) and Claudia Marsicano (Universidad de Buenos Aires in Argentina) both joined the temnospondyl efforts starting in the 80s and 90s (arguably the majority of their work and their most prominent publications have been in the 2000s).
Representation from outside of Europe/North America and among women *has* improved in the 21st century - researchers such as Graciela Piñeiro (Universidad de la República), Dorota Konietzko-Meier (Opole University), and Sophie Sanchez (Uppsala University) are some of the most established female temnospondyl workers; Hillary Maddin (Carleton University) and Nadia Fröbisch (Museum für Naturkunde) also have a great track record, although they're both trending more toward evo-devo these days. Julia McHugh (Museums of Western Colorado) and Kat Pawley (formerly LaTrobe) both did their dissertations on temnospondyls but Julia hasn't published on temnos in a few years, and Kat is out of the field now. We're also starting to see some homegrown researchers outside of the Euramerica complex, particularly as temno research is picking up in South America; Estevan Eltink (Universidade Federal do Vale do São Francisco in Brazil), Sérgio Dias-da-Silva (Universidade Federal de Santa Maria in Brazil), and Dhurjati Sengupta (Indian Statistical Institute) are three who immediately come to mind. My buddy Calvin So (PhD student at George Washington), Ben Otoo (PhD student at UChicago), Elzbieta Teschner (Opole University), and Sanjukta Chakravorti (recent PhD grad at the Indian Statistical Institute) are some up-and-coming women and/or POC to keep an eye on, and please look for ways to be inclusive in your temno research teams!
8. Almost 200 years of temnospondyl research and there's still zero consistency in what to call enlarged palatal teeth in temnospondyls.
Fangs? Tusk? "Fangs?" "Tusks?" I've already blogged about this before, but it sure it irritating (not that I'm any better at maintaining consistency) to have a bunch of terms, really none of which are appropriate in a strict sense anyway, floating around in the literature. Just one of those pet peeves things that make academics special... [Zach Lowe often has one short toss-away in his "10 Things," so this is my equivalent]
9. Jurassic World - The Game depictions of temnospondyls.
Temnospondyls haven't made it into a documentary since BBC's 2005 documentary "Walking with Monsters" in which an aestivating Rhinesuchus got eviscerated by a giant gorgonopsid (it's episode 6 if you want to watch the horror). They didn't make it into the Jurassic Park reboot either, but they did make it into the 2015 mobile game, the one that's like zoo tycoon, not the one where you use a drone to somehow collect DNA from AR dinosaurs and that destroys your battery and your data plan simultaneously. There are a bunch of probable temnospondyls - Koolasuchus, Microposaurus, 'Labyrinthodontia,' which I can only assume is supposed to be Mastodonsaurus (but also Mastodonsaurus is its own thing), Metoposaurus (!), Prionosuchus . Since Mastodonsaurus is an actual animal, it's not clear what 'Labyrinthodontia' is supposed to be? But it's in the "Common" tier, so evidently not a very good one. The weird little factoid for the first level states: "Did you know that it's believed that the Labyrinthodontia is the progenitor of all modern amphibians?" I'm pleasantly surprised that Metoposaurus is not only in the "Legendary" tier (the highest one) but can also be collected only through VIP packs (pay-to-play).
All things considered, the base models look okay. Mastodonsaurus....something weird's going on with that dude's eyes, and Microposaurus' head just doesn't look right. But other than that, proportions and anatomy aren't totally off-base. It's when you get to the "evolved" (powered-up) forms that things get really weird...whoever designed this game decided that "evolution" meant you got amazingly flamboyant, and as far as the temnospondyls go, grew some wild dorsal fins! The one on Prionosuchus is particularly...magical.
On the off-chance that anyone from Ludia is reading this blog...I can help you fix your temnospondyls!
10. Newly outdated signage in newly renovated exhibits.
Nothing says "new year" like new exhibits with signs that are already out of date (the revamped University of Michigan Natural History Museum is amazing in spite of this - check it out for sure!).
POWER RANKINGS
As some people know, I am a HUGE basketball fan as well (Laker fan for life!), and I am an avid reader of the regular NBA power rankings that are put out by ESPN. These are almost entirely arbitrary rankings (although this is not a "list of Bryan's favourite and least favourite temnospondyls") and are based on zero quantitative metrics beyond my own perceptions of the body of the work that's been put out there, but it did add up to a nice, round 20 clades.
Stars (Lebron James tier)1. Capitosaurs
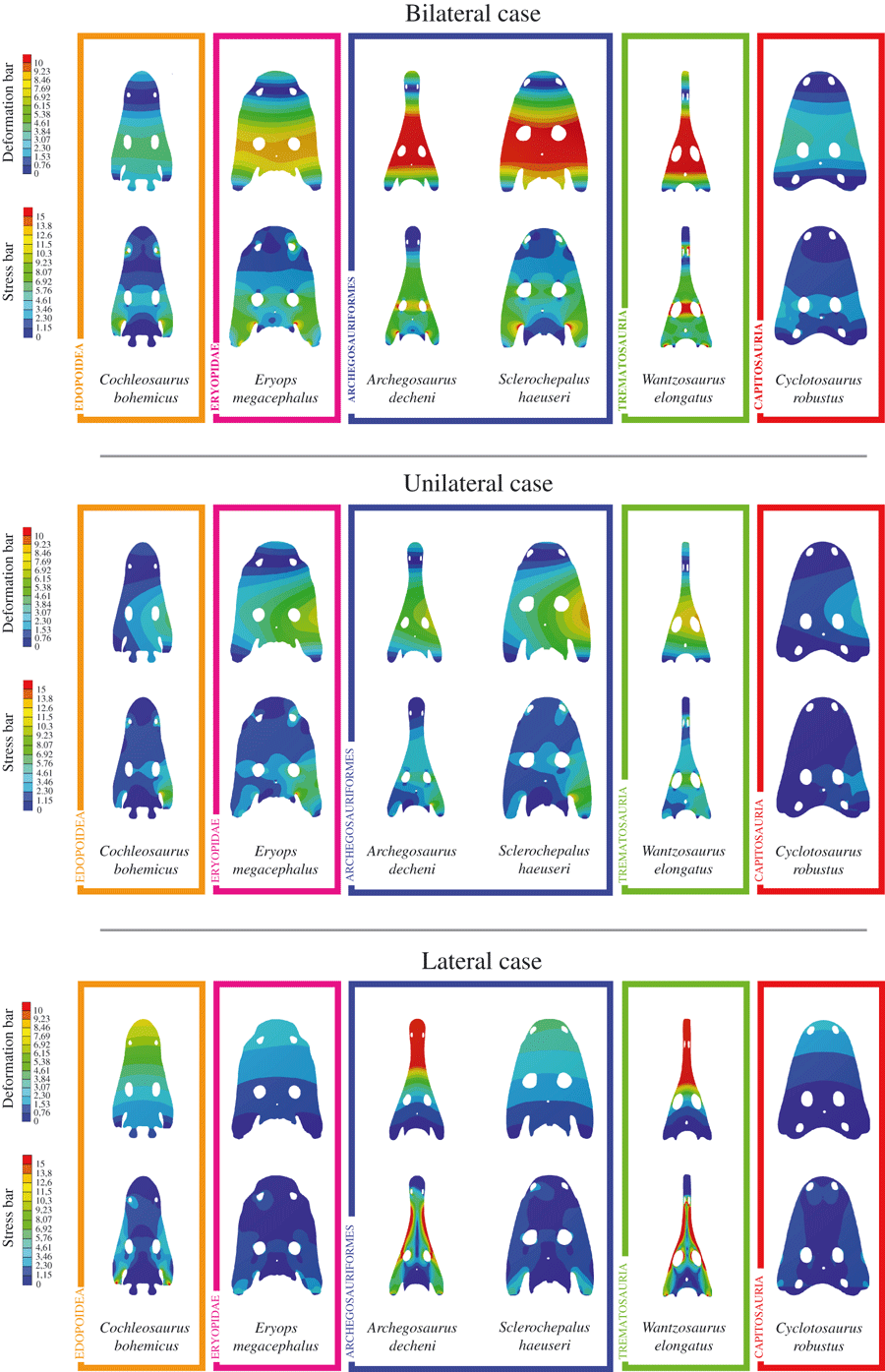
Nothing says 'dynasty' like including the first temnospondyl to be named (Mastodonsaurus) and then still being so beloved that there are more than a dozen papers on your clade. Capitosaurs have had more new taxa described than in any other temno clade, and it's not even close; in the 2010s, we got Antarctosuchus polyodon Sidor, Steyer, & Hammer, 2014; Calmasuchus acri Fortuny, Galobart, & De Santisteban, 2011; Cyclotosaurus buechneri Witzmann, Sachs, & Nyhuis, 2016; Eocyclotosaurus appetolatus Rinehart, Lucas, & Schoch, 2015; Parotosuchus ptaszynskii Sulej & Niedzwiedzki, 2013; Samarabatrachus bjerringi Novikov, 2016; Selenocara rossica Novikov, 2016; Stanocephalus amenasensis Dahoume et al., 2016; Syrtosuchus morkovini Novikov, 2016; Tomeia witecki Eltink, Stock Da-Rosa, & Dias-da-Silva, 2016; and Yuanansuchus maopingchangensis Liu, 2016. Additionally, there was the first report of any temnospondyl (a capitosaur) from Japan (Nakajima & Schoch, 2011), further description of material of E. appetolatus (Rinehart & Lucas, 2016), revision of capitosaur material from the Arctic (Kear et al., 2016), and revision of the well-known Parotuschus nasutus (Schoch, 2018). Capitosaurs were also the "test subjects" for a lot of the finite-element analysis (FEA) work done by Fortuny and colleagues (Fortuny et al., 2012, 2016; Marcé-Nogué et al., 2015), and some recent histo work suggested that some might have been more terrestrially capable than traditionally conceived (Mukherjee et al., 2019).
2. Metoposaurids
Sidekicks (CJ McCollum tier)3. Amphibamiforms
Some name changes and their perennial status as probable stem-lissamphibians keep amphibamiforms in the spotlight. For starters, the clade traditionally termed 'Amphibamidae' that encompassed small-bodied terrestrial dissorophoids is now restricted to just Amphibamus and Doleserpeton (Schoch, 2019a); 'Amphibamiformes' now encompasses all of the traditional amphibamids and branchiosaurids. There were a few new taxa: Rubeostratilia texensis Bourget & Anderson, 2011; Nanobamus macrorhinus Schoch & Milner, 2014 (then re-described by Gee & Reisz, 2019); Platyrhinops fritschi Werneburg, 2012; and Tersomius dolesensis Anderson & Bolt, 2013; as well as a thorough redescription of Doleserpeton annectens (Sigurdsen & Bolt, 2010), revisions of Tersomius texensis (Maddin et al., 2013), Milnererpeton huberi (Werneburg et al., 2013), Branchiosaurus salamandroides (Werneburg, 2012) and additional reports of Leptorophus (Schoch, 2014a). Werneburg (2019) revised a number of Permo-Carboniferous specimens from France, erecting the terrestrial amphibamiforms Anthracobamus fayoli and Montceaubatrachus platyrynchus and a branchiosaurid Branchiosaurus commentryensis. A complete but badly weathered branchiosaurid was also reported from Turkey by Fortuny et al. (2015). There were three great histological studies by Sanchez et al. (2010a, 2010b, 2010d) that used Apateon to examine different paleoecological and paleoenvironmental attributes, and Eichler & Werneburg (2010) examined a number of branchiosaurids using UV light. Schneider & Werneburg (2012) provided an updated stratigraphic framework of the Rotliegend branchiosaurids. Even if they prove to not be ancestral to all modern amphibians, their diminutive body size (referred to by some as 'miniaturization'), extensive sample size for some taxa, and exceptional preservation of soft tissue will keep them in the limelight for a long time.
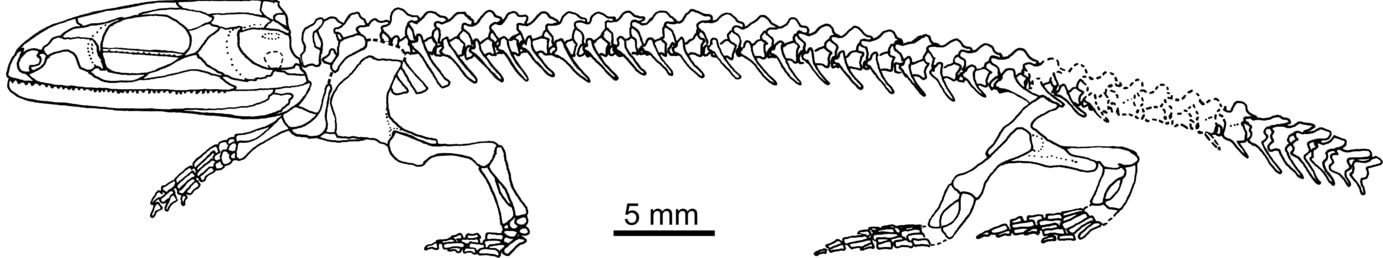
Reconstruction of Doleserpeton annectens from the early Permian of Oklahoma (figure from Sigurdsen & Bolt, 2010).
5. Dissorophids
Dissorophids have had one heck of a renaissance in the 21st century. In the last decade, there have been five new taxa (Broiliellus reiszi Holmes, Berman, & Anderson, 2013; Cacops woehri Fröbisch & Reisz, 2012; Nooxobeia gracilis Gee, Scott, & Reisz, 2018; Reiszerpeton renascentis Maddin, Fröbisch, Evans, & Milner, 2013; and Scapanops neglecta Schoch & Sues, 2013), as well as the first phylogenetic analysis of the clade (Schoch, 2012), detailed descriptions of several eucacopines (Gee & Reisz, 2018; Liu, 2018), and some promising reports of Carboniferous-age material from the midcontinent of North America (May et al., 2011). Numerous sources indicate exciting forthcoming work from areas outside of the classic collecting localities for the clade...

Three specimens of Cacops morrisi from the early Permian of Oklahoma (figure from Gee & Reisz, 2018).
Sixth men (Lou Williams tier)
8. Rhinesuchids
This clade lives and dies by the Brazilian rhinesuchid Australerpeton cosgriffi, which has been a real stand-out among the rhinesuchids, with a cranial osteology packaged in with a phylogenetic analysis of Rhinesuchidae (Eltink et al., 2016) and two reports of new specimens (Eltink & Langer, 2014; Azevedo et al., 2019). A second phylogenetic analysis was published by Marsicano et al. (2017) just a short while later. Piñeiro et al. (2012) described a new taxon from Uruguay, Arachana nigra, and there are also promising reports of indeterminate rhinesuchids among the early Permian Brazilian assemblages (Cisneros et al., 2015). McHugh (2014) sliced and diced the eponymous Rhinesuchus. Arguably the ace-in-the-hole is the poorly known Broomistega, an individual of which was found cuddling a therapsid in a burrow (Fernandez et al., 2013).
Role players (PJ Tucker tier)10. Eryopoids
The most iconic Paleozoic temnospondyl (and arguably, tetrapod, other than Dimetrodon) might never have a heyday as great as the early 20th century when everybody and their mother (or father, let's be real) published papers about every specific region of the skeleton of Eryops. The 21st century has mostly seen simple reports / documentation of various taxa (mostly Eryops) from different parts of the world (mostly North America) (e.g., Werneburg et al., 2010; Witzmann, 2013; Aber et al., 2014; Voigt et al., 2014; Witzmann & Voigt, 2015; Huttenlocker et al., 2018). Keep your eyes peeled for a new species of Eryops though (Rasmussen et al., 2016)! On the descriptive side, Čerňanský et al. (2016) published a CT analysis of the quadratojugal of Eryops, and Werneburg & Berman (2012) touched up the osteology of Glaukerpeton avinoffi.
12. Stereospondylomorphs (in the paraphyletic sense)
A pretty low-key decade for the early stereospondylomorphs (the non-stereospondyl stereospondylomorphs to be technical). A few new taxa popped up as well, such as Konzhukovia sangabrielensis Pacheco, Eltink, Müller, & Dias-da-Silva, 2017 and Sclerocephalus stambergi Klembara & Steyer, 2012. The holotype of the type species of Sclerocephalus, S. haeuseri, which was long thought to be lost, also reappeared this decade (Schoch et al., 2019). Stereospondylomorphs also got a brand new phylogenetic analysis (Eltink et al., 2019), with the erection of a few new clades. Witzmann & Brainerd (2017) modeled the physiology of Archegosaurus decheni. Like dvinosaurs, the early stereospondylomorph record from South America is developing really quickly and in very interesting ways, with a lot of connections to eastern Eurasia (particularly Russia) that are changing a lot of the ways in which we historically conceived of biogeographic distribution and radiation of stereospondylomorphs.
13. Micromelerpetids
Like rhinesuchids, micromelerpetid research rests largely on the eponymous Micromelerpeton; the only other major descriptive work is the first report of the clade in Africa (Werneburg et al., 2019) and a re-description of Limnogyrinus with a phylogenetic analysis (Schoch & Witzmann, 2018). Unlike rhinesuchids, there's no micromelerpetid cozying up with an amniote. BUT, micromelerpetids do show evidence of limb regeneration and preaxial polarity during limb formation (Frobisch et al., 2014, 2015), which makes them super interesting from the evo-devo side of things.
Riding the bench ("pick any obsolete rookie" tier)14. Brachyopoids
The two brachyopoid clades, Brachyopidae and Chigutisauridae, have been pretty meager. The main work on the brachyopid front was Warren et al.'s (2011) documentation of tupilakosaur-like vertebrae (better known as the diplospondylous type) in Bothriceps. Chigutisaurids faired a little better. Compsocerops, originally described from India, was found in Brazil (Dias-da-Silva et al., 2012). Some chigutisaurid material from India was also sampled in a comparative histological analysis (Muhkerjee et al., 2010); word on the street is there's more histo coming out soon! There's hope on the horizon for brachyopoids if future work bears out their relationship to 'latiscopids,' and in turn, 'latiscopids'' close relationship to caecilians.
15. Rhytidosteids
Next to nothing on these small-bodied and typically horned temnos, which occupy an important position in the context of the early radiation of Mesozoic stereospondyls. Dias-da-Silva & Marsicano (2011) did provide the most comprehensive phylogenetic analysis of the group to date, although many of their results were challenged by Maganuco et al. (2014) who re-described Mahavisaurus dentatus. Sennikov & Novikov (2018) discussed possible trophic adaptations / specialization for hard-bodied invertebrates.
16. Edopoids
Apparently little appetite for the "gluttonous faces" of the Paleozoic. Essentially no descriptive work beyond some new material of the relict Nigerpeton (Sidor, 2013) and only brief discussion in broader comparative papers (e.g., Fortuny et al., 2011; Schoch, 2019b). Bird et al. (2019) did squeeze a paper on a trackway attributed to an edopoid from the lower Carboniferous of the UK into 2019.
17. Dendrerpetids
Other than having the honour of being utilized as the outgroup for just about every Paleozoic temnospondyl study, dendrerpetids have been on the down and out for some time. New material and dedicated re-analysis to the ages-old question of "just how many dendrerpetids are there at Joggins" may inspire some work going forward (Maddin et al., 2018).
18. Lydekkerinids
What happens when you're small, fairly abundant, not closely related to modern amphibians, and phylogenetically well-resolved? You guessed it - you get sliced and diced (Canoville & Chinsamy, 2015; McHugh, 2015) and that's about it...
19. Zatracheidids
Does anybody still work on zatracheidids? Zero publications on this clade in the 2010s aside from their sporadic inclusion in broader analyses of temnospondyl macroevolutionary patterns (e.g., Angielczyk & Ruta, 2012; Witzmann & Ruta, 2019).
20. Lapillopsids
Guess what happens when you're small, one-of-a-kind, and a phylogenetic wildcard? You guessed it - no research in the 2010's! In fact, you only get six hits on Google Scholar from 2010 onward.
Five predictions for the 2020's
I will freely admit that I am totally spitballing here, but it'll be interesting to see how these pan out.
Five things I wish for but don't reasonably expect in the 2020's
Social Media Round-up
(Long list of references for this post, so I've hidden them below the "Read More" line). Note that this is not at all an exhaustive list of all temnospondyl publications since 2010; I haven't even cited some of my own here.
Temnospondyls are undoubtedly some wonky animals in general, as they look like a cross between a salamander and a crocodile and do all sorts of weird and wonderful things. Ned Colbert once referred to stereospondyls as "queerly proportioned beasts such as we might encounter in some mediaeval manuscripts," which is an excellent tagline for a LinkedIn page. Zatracheidids, though not stereospondyls, are "queerly proportioned" so to speak and have also inspired some really great quotes, perhaps not quite at the typical level of proper academic writing, that live in perpetuity in the literature. Below are some of my favourites:
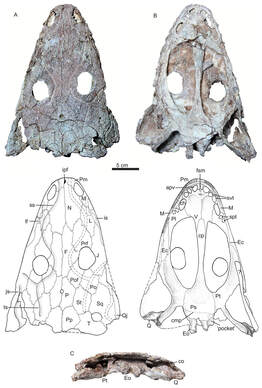
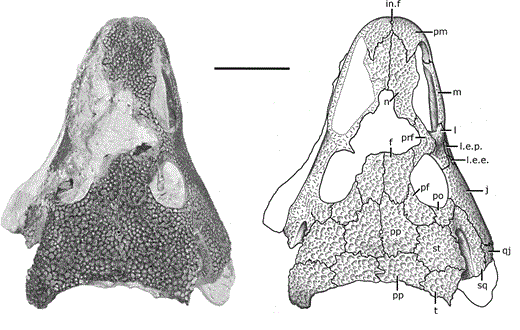
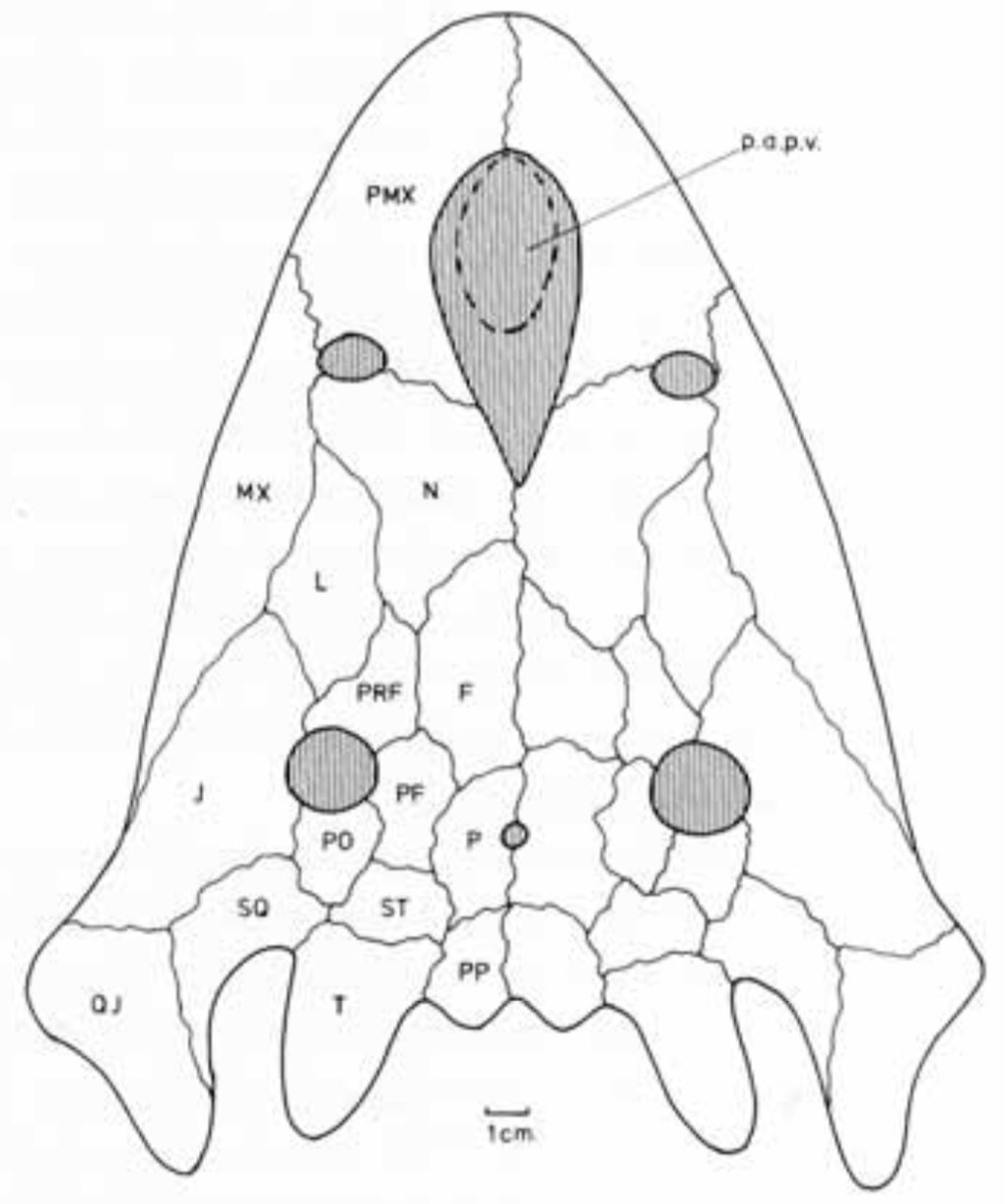
AcanthostomatopsAcanthostomatops is the first of two zatracheidids from Europe, being known from several dozen specimens collected from an early Permian locality near Dresden, Germany. Of the zatracheidids, it is the smallest, has the proportionately widest skull and smallest internarial fontanelle, and the best-known ontogenetic series (see below), described both by Boy (1989) and Witzmann & Schoch (2006). A larval specimen reported by Werneburg (1998) has been re-identified as a micromelerpetid by Witzmann & Schoch (2006). The genus was originally named Acanthostoma, with detailed descriptions being made by Credner (1883) and Steen & Brough (1937), but this name was already taken by some esoteric polychaete worm that got the name 70 years before the temnospondyl, so it changed in 1961 by German paleontologist Oskar Kuhn. Among the detailed ontogenetic information available through the large sample size is an apparent change in the hyobranchial apparatus from larval forms to adults. Witzmann & Schoch (2006) suggested that this might relate to the development of a complex tongue apparatus that would have been adaptive in terrestrial environments, possibly as an ambush predator. 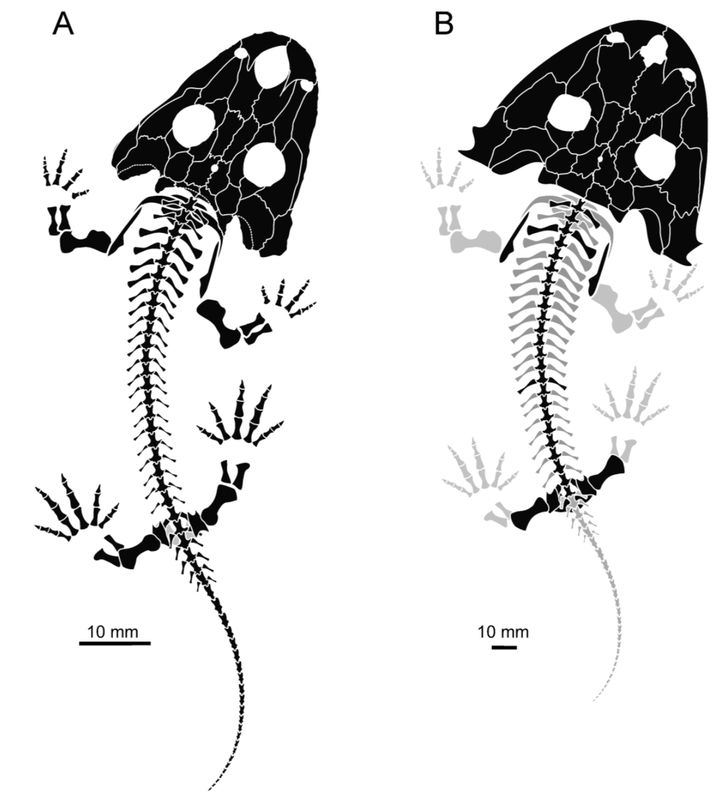 DasycepsDasyceps is one of the few Paleozoic temnospondyls from the U.K. It was originally described as a species of the genus "Labyrinthodon" (no longer valid, now a junior synonym of Mastodonsaurus) but was subsequently recognized as a distinct species by Thomas Huxley in 1859 and given its present name. It was redescribed (in German) by von Huene in 1910, and following the description of the other zatracheidids, also identified as such early in the 20th century by Case. At one point, it was regarded as synonymous with Zatrachys (Broom, 1913), but this opinion never really took off, and the possible juvenile-adult relationship between Acanthostomatops and Zatrachys has garnered more attention in recent history. Dasyceps is the largest known zatracheidid, with a skull length approaching 30 cm, and the skull is proportionately long compared to the shorter and more parabolic skulls of Acanthostomatops and Zatrachys. Its internarial fontanelle is a perfect teardrop and is preserved as such in the holotype - it's not a reconstruction that is artificially smoothed to look nice. Other features in which it differs from other zatracheidids include a narrower otic notch, relatively smaller orbits and nares, and broad, posteriorly extensive tabular horns (slender in Zatrachys). It also lacks the spiky projections from the lower cheek region that are found in other taxa, although it still has markedly expanded quadratojugals. Whether some of these features are associated with size (and possibly advanced ontogenetic maturity) is unclear.
The first species of Dasyceps is Dasyceps bucklandi, named after the famous 19th century paleontologist William Buckland. A second species from the early Permian of Texas, Dasyceps microphthalmus, was originally described in 1896 by Cope as a species of Zatrachys; it was subsequently moved over by Paton (1975). This species is much more poorly known than D. bucklandi, being represented only by a poorly preserved holotype "covered in bean ore" (Cope, 1896), whatever 'bean ore' is, and two fragmentary referred skulls. If the generic assignment is correct (I have no reason to question it), this would be one of the most geographically wide-ranging genera of Paleozoic temnospondyls (ignoring the present-day separation of the U.K. and Texas); most taxa, particularly terrestrial ones, are fairly restricted. Milner et al. (2007) identified a small zatracheidid from the early Permian of the Czech Republic that they referred to Dasyceps sp. ZatrachysZatrachys is one of the more iconic North American Permian temnospondyls if only because zatracheidids generally look weird. The first species, Z. serratus, was named in 1878 by E.D. Cope for material collected from Texas, but the material was relatively fragmentary, which led to reconstructions that more closely resembled Dasyceps. Additional, seemingly by random chance, the holotypes of Z. serratus and 'Z. microphthlamus' (now in Dasyceps) are lost, as is that of 'Zatrachys conchigerus,' which is now a nomen vanum. None was ever very clearly illustrated either. The neotype of Z. serratus is just the back half of a skull, which is badly weathered, thus preserving little of the cheek spines and none of the internarial fontanelle. The best-known material is thus that described from New Mexico by Langston (1953) and Schoch (1997), which includes complete skulls of several growth stages and which captures a range of intraspecific variation. A number of other species have been named under Zatrachys, but most of these are now regarded either as indeterminate temnospondyls or more specifically as some kind of aspidosaurine dissorophid, and it has also been confused with the dissorophid Platyhystrix on account of erroneous association of skeletal remains. Among them are Zatrachys apicalis and Zatrachys crucifer, which are both placed in Aspidosaurus now. 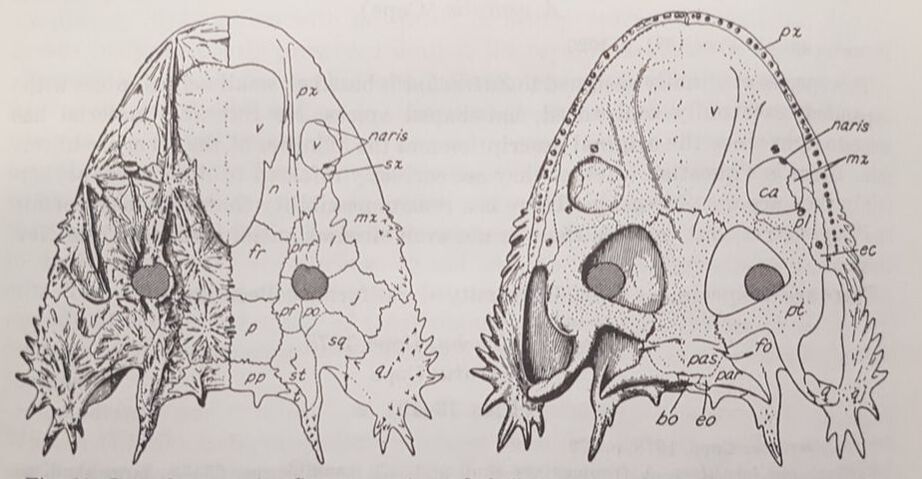 Reconstructions of the skull of Zatrachys serratus (figure from Langston, 1953).  Photo of the Pinocchio Frog; © Tim Laman/National Geographic Photo of the Pinocchio Frog; © Tim Laman/National Geographic So what's the giant hole for? The popular suggestion of the time, which is arguably still the popular suggestion of today, is that the fontanelle of temnospondyls housed a gland at least similar to, if not homologous with, the glandula intermaxillaris in modern amphibians (e.g., von Huene, 1910; Säve-Söderbergh, 1935), which is what produces the sticky secretion that then gets adhered to the tongue for snagging insects. This does explain why many terrestrial temnospondyls have a fontanelle, with a corresponding intervomerine depression just below it on the roof of the mouth, but it fails to account for the massively enlarged fontanelle of zatracheidids - what would compel an animal to need so much of that secretion compared to other animals of a presumed similar ecology? It's not even clear, as noted in the above quote by Paton (1975), whether Dasyceps could sustain itself on insects given its size (though sticky secretions can probably help snag small tetrapods too). If the gland was indeed much larger and produced a much larger volume of secretion, then it would probably have to be a very situational thing, perhaps akin to how hagfish exude thick coats of mucus to clog a predator's mouth, otherwise the amount of water required to produce that much mucus would be prohibitively costly for the animal. So the TLDR is that it doesn't seem very likely that the gland would be orders of magnitude larger than in any other temnospondyl. The better, although hardly substantiable, suggestion is that of Paton (1975), who provided a detailed re-description of Dasyceps from England. She suggested that the fontanelle in zatracheidids housed an expandable sac that would inflate as a defence mechanism. There is admittedly no real justification for this, but it does vaguely remind me of the Pinocchio Frog (discovered in 2008, described in 2019), which can inflate that little trunk it's got sticking off its face. What about the spikes? Other than Langston's (1953) suggestion that the spikes made Zatrachys less palatable to sharks, there isn't a clear function of the spikes, which are also absent in Dasyceps. Langston compared them to those of phrynosomatids, more commonly called 'horned toads' (but they're lizards). There is also an appreciable amount of variability in the development of the spines within Zatrachys; they may have only been ornamental structures. The presence of spikes in the Carboniferous-age Stegops has long been cited as evidence for affinities of that taxon with zatracheidids, but it more commonly comes out as an early dissorophoid. Updated Wikipedia articles! I don't publicize this much, but I go through from time to time, usually when I'm procrastinating my dissertation, and touch up (i.e. massively expand) the articles for early tetrapods that I have some familiarity with (I actually don't do metoposaurids much because of a concern, probably unfounded, that I'm too close to the subject matter to be unbiased in how I present certain information). Most of the "lower tetrapods" only have a single sentence, so there's a lot of work to be done. That was the case for all of the zatracheidids, so I've updated those pages along the lines of the content I wrote above: Refs
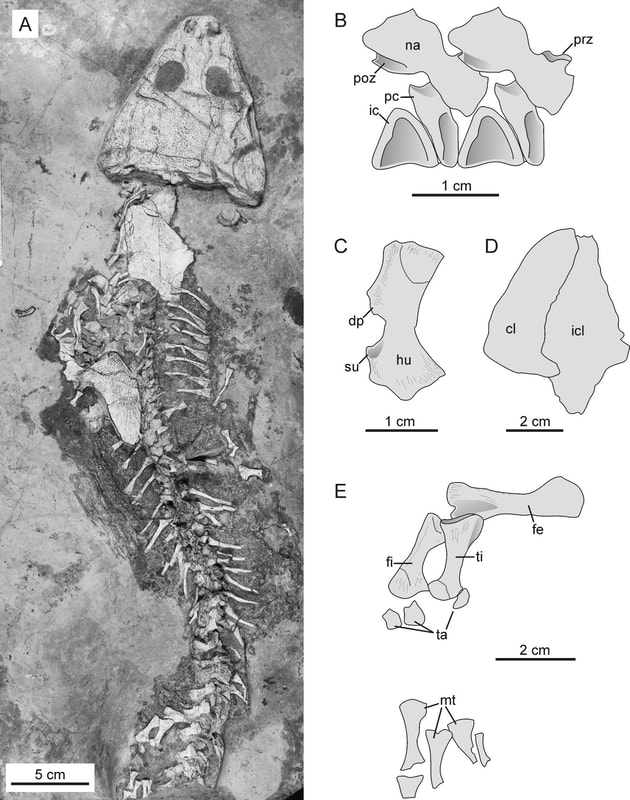
Fresh off a collections visit to the Carnegie Museum in Pittsburgh, it seems only fitting to blog about what I think is one of the greatest strengths of their collection - the Paleozoic tetrapods! The Carnegie owns one of the largest collections of Paleozoic tetrapods from the American southwest (especially New Mexico) and from the more geographically proximal Dunkard Basin (Ohio, Pennsylvania, West Virginia). Much of this was both collected and studied by emeritus curator David Berman and collections manager Amy Henrici, as well as the late Peter Vaughn, a longtime professor at UCLA who taught Dave (among many other paleontologists). Among the stash of early tetrapods are important holotypes of three trematopids, the large dissorophoids with peculiarly long nostrils that I've done some work on myself, so it seems only fitting to do a blog post highlighting a little bit more about these weird animals.
|
About the blogA blog on all things temnospondyl written by someone who spends too much time thinking about them. Covers all aspects of temnospondyl paleobiology and ongoing research (not just mine). Categories
All
Archives
January 2024
|














 RSS Feed
RSS Feed
