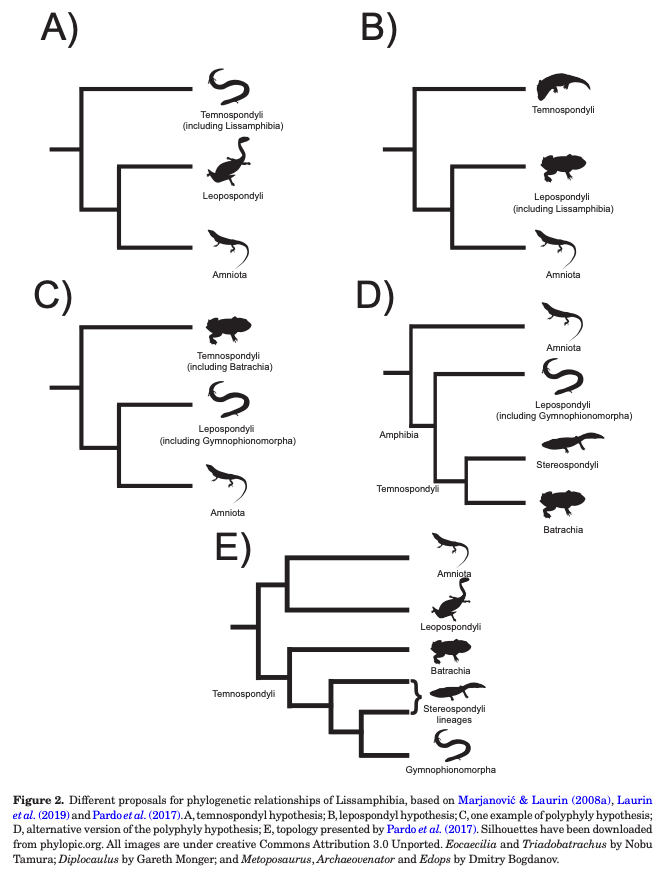
|
2020 has certainly been some kind of year, which makes it easy to miss the latest scientific discoveries related to everyone's favourite (usually) four-fingered tetrapods. Last year I did a wrap on the entire decade, so there was a quite a lot, but in spite of the circumstances, 2020 has still produced some very exciting temnospondyl research! As with last year's wrap-up, this focuses on temnospondyl-centric research; there are obviously plenty of papers that make cursory mention of them or that might include a picture or two in an assemblage description, but those are not summarized here (I gotta be efficient with my time). As usual, links to everything are in the reference list as the end. Hopefully looking forward to getting back on a more regular track again in 2021 - stay tuned! Leaving a markIn lieu of body fossils, we often have evidence of temnospondyls in the form of trace fossils, which usually consist of a trackway. In contrast to amniotes, which basically all have pentadactyl (5-digit) hands, temnospondyl trackways have been considered to be historically easy to ID because of the tetradactyl (4-digit) hand that typifies temnospondyls and many modern amphibians (at least the ones that still have hands). Body and trace fossils rarely mix because the optimal conditions to preserve both the hard parts and the traces of activity will differ somewhat starkly, but this is good because it means where we might lack a body fossil record, perhaps we could supplement the total record with traces.
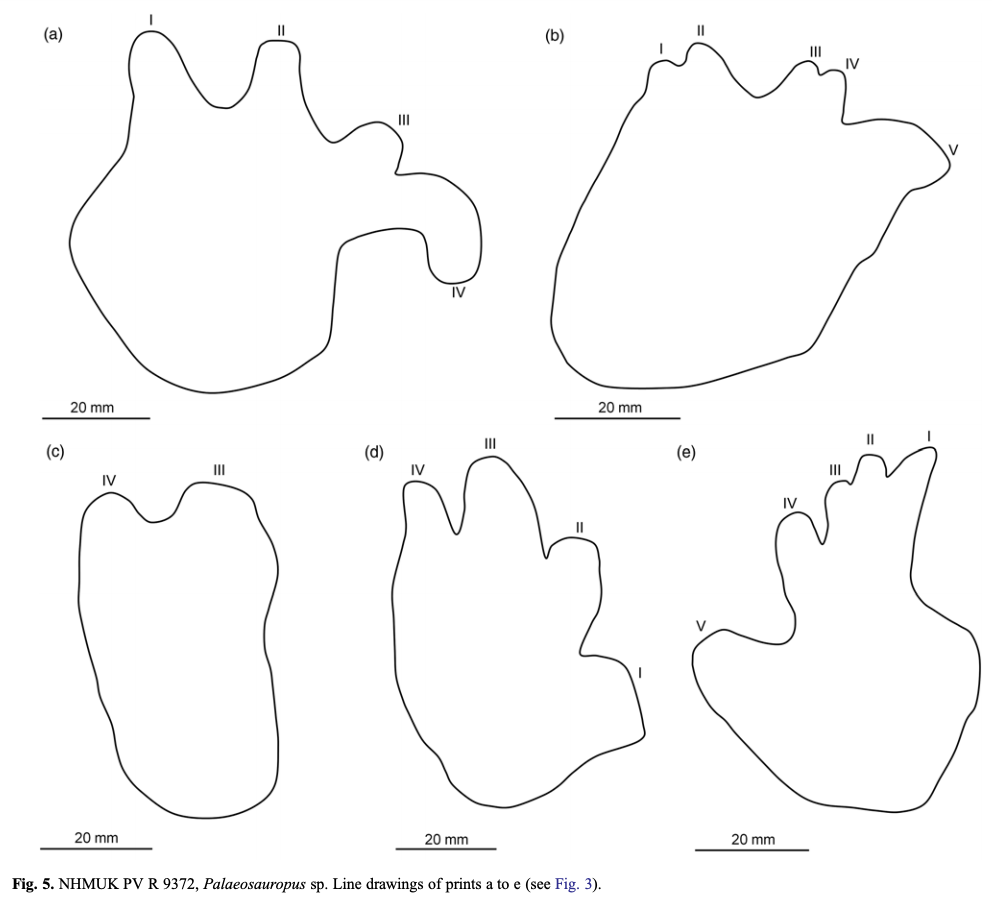
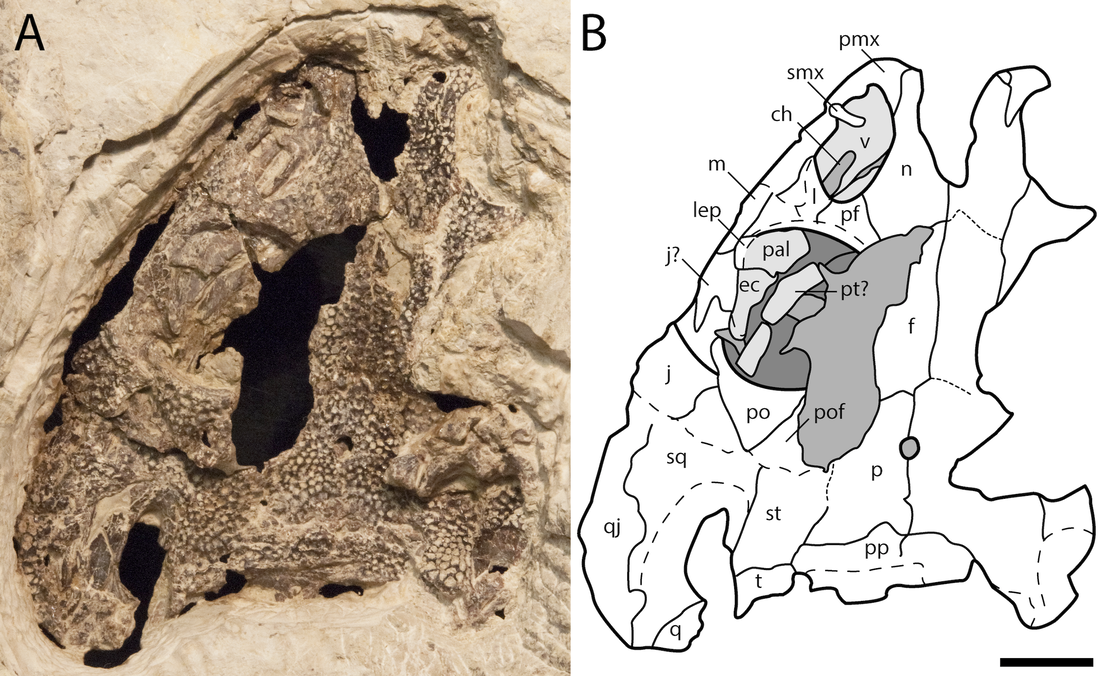
Bird et al. (J Geol Soc) reported a trackway from the early Carboniferous (Visean) of the U.K. (paper technically online in late December last year, but in print this year). This time period is a critical window of tetrapod evolution, when stem tetrapods really radiate; however, we still don't have most of the groups that directly lead to modern groups, and there is only one Visean temnospondyl, Balanerpeton woodi from Scotland (which is also debated about whether it might be a stem tetrapod and not a temnospondyl). The U.K. in general is pretty sparse on the Paleozoic tetrapods. As such, this is a good example where the trace fossil record pre-dates the main body fossil record and thus hints at a large gap in the body fossil record of the earliest stages of a group's evolution (temnospondyls in this case). The ichnotaxon (taxonomic framework for trace fossils) that they assigned the material to is Palaeosauropus, which is also known from the Blue Beach locality in Nova Scotia and Pennsylvania and which was previously interpreted to belong to an edopoid (the validity of this association is a little questionable). The oldest body fossil record of an edopoid is the Bashkirian, which begins around 323 million years ago, whereas the Visean ends at 330 million years, so at minimum, this possible edopoid record extends the group back by at least 7 million years, which is congruent with phylogenetic analyses that recover edopoids as a very early diverging group.
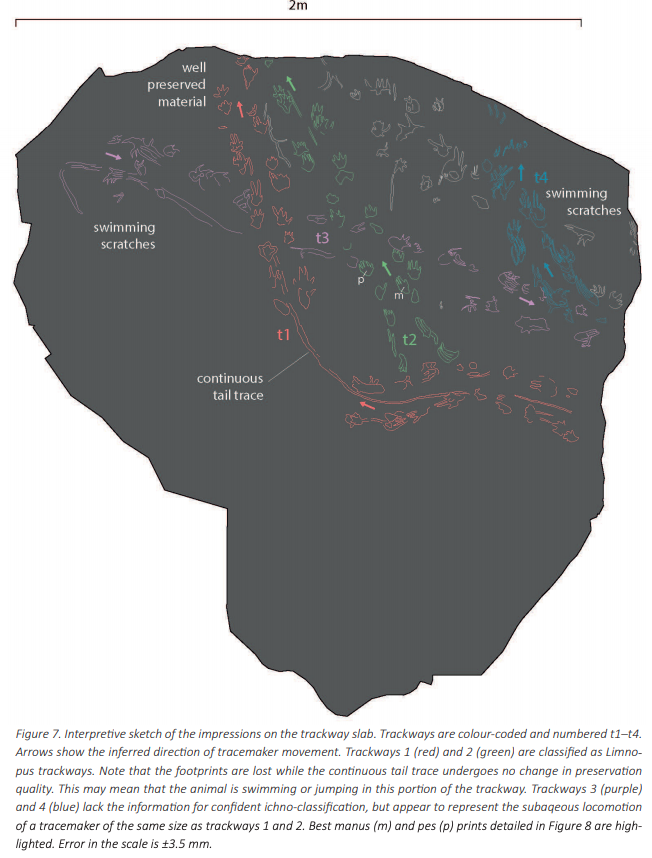
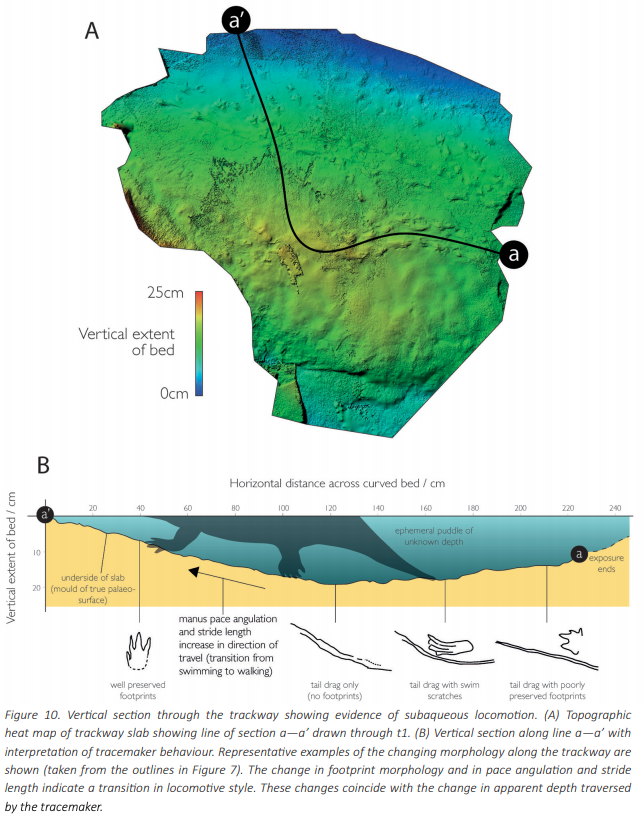
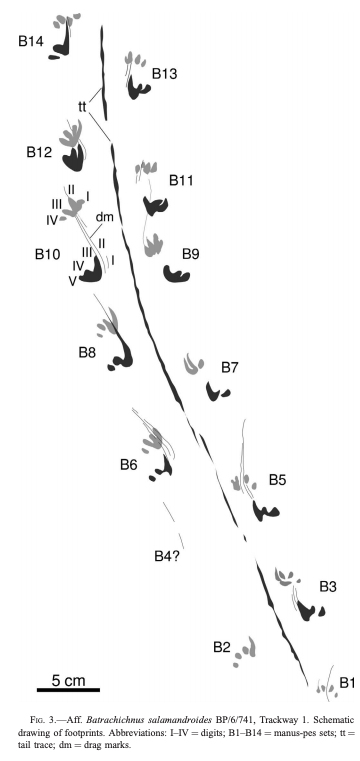
Herron et al. (Norwegian J Geol) reported slightly younger tracks from the Late Carboniferous (Moscovian-Kasimovian), when the temnospondyl body fossil record is much better established, from Svalbard. This archipelago is usually better known for its Triassic temnospondyl body fossil, which compares favourably with the closely situated Greenland assemblage, and the Scandinavian Arctic region has long been productive for some of the most important stem tetrapod fossils like Acanthostega. The new material that Herron et al. report is a massive block (almost 800 photos to produce a composite model) that captures a rare setting: the margin of a pond. As a result, the trackway captures the transition between the animal moving in the deeper part of the pond (with more scratch-like traces made through minimal contact with the pond bottom) and then moving out onto land (with more pronounced footprints).
There were a few other papers that briefly show some temnospondyl traces or trackways that might be temnospondyls:
An abundance of skepticism
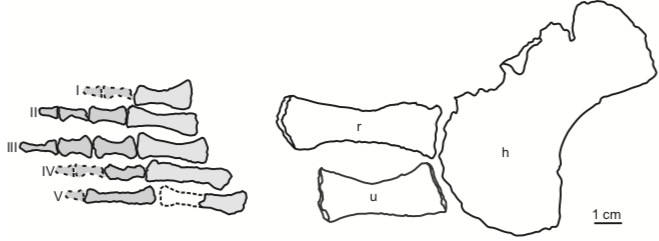
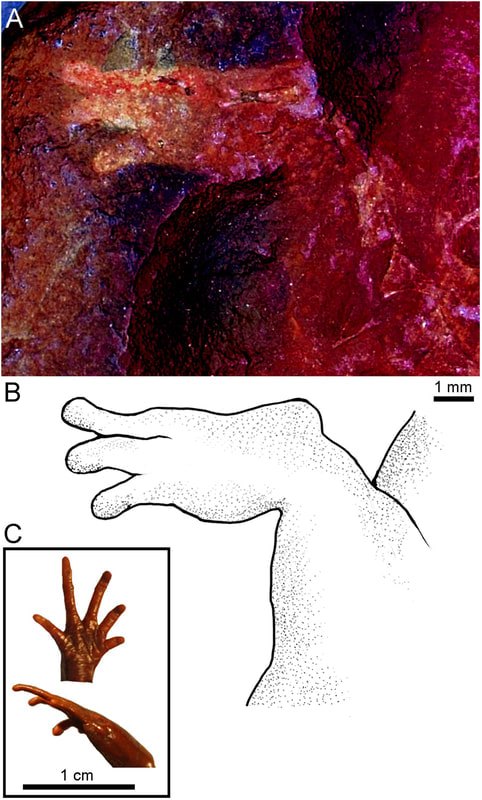
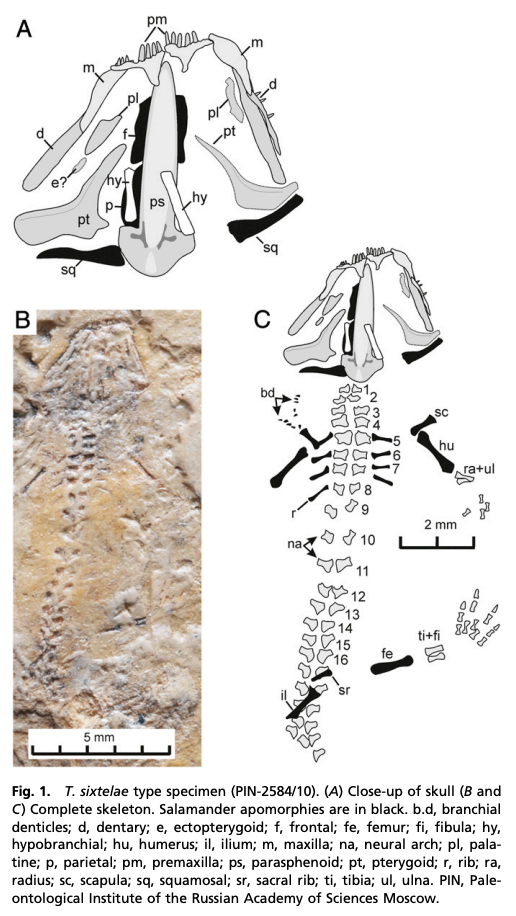
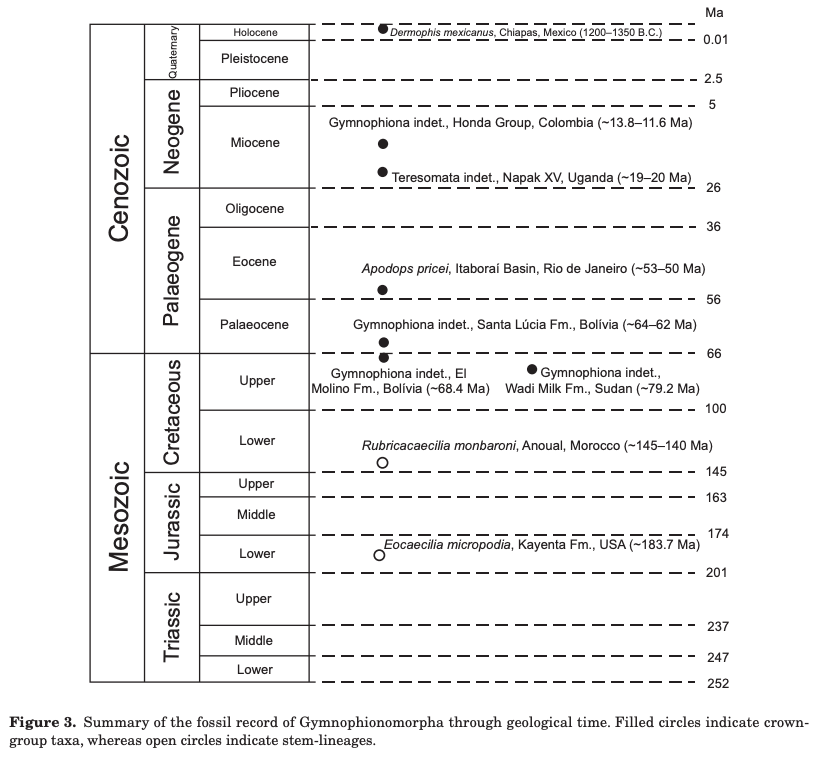
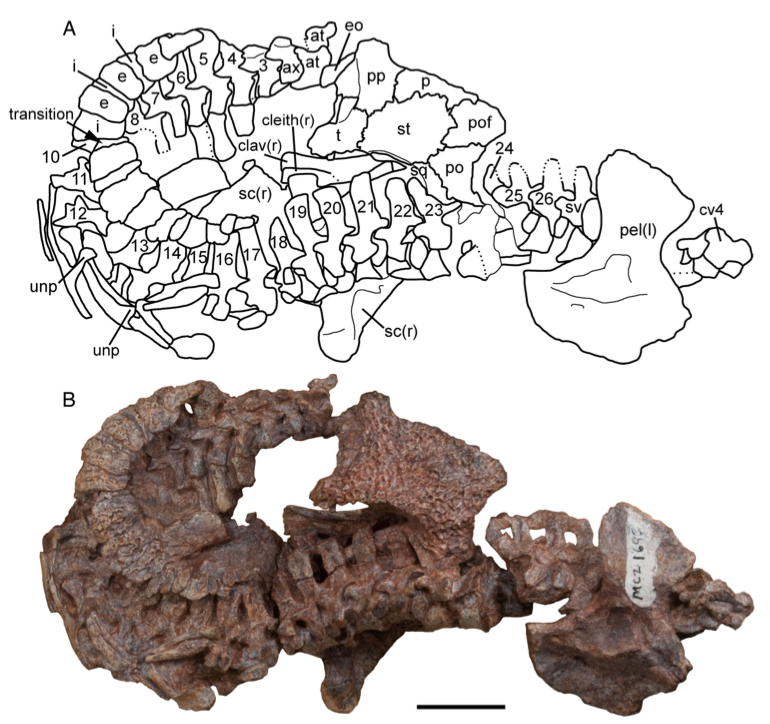
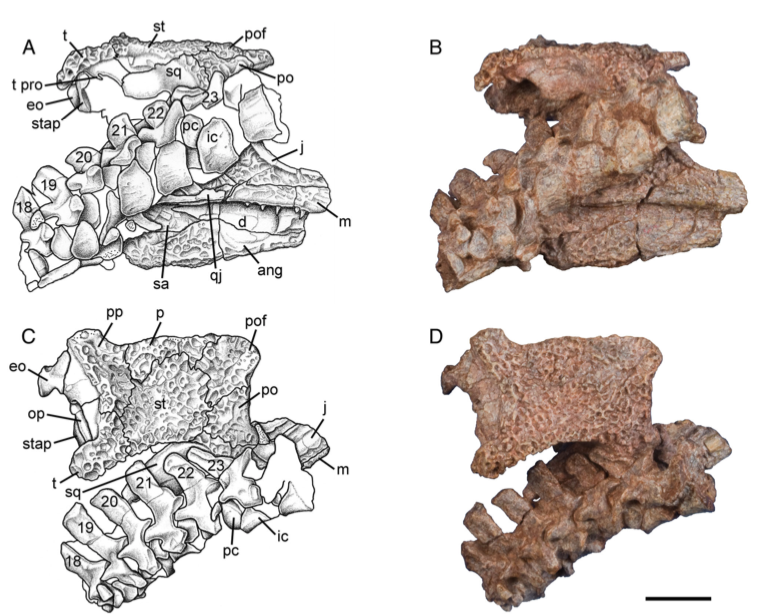
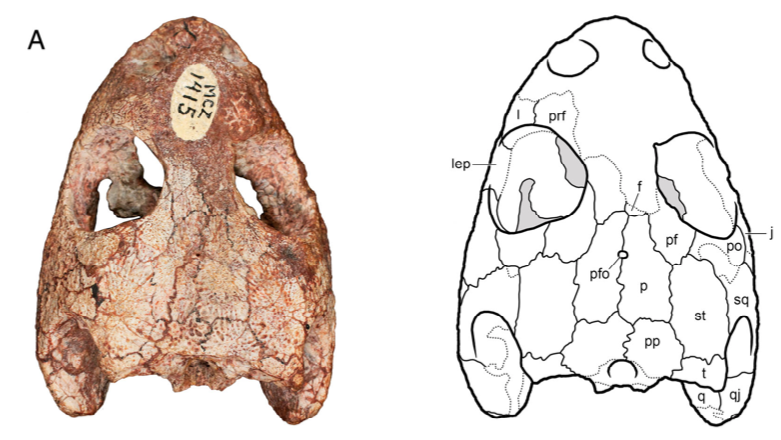
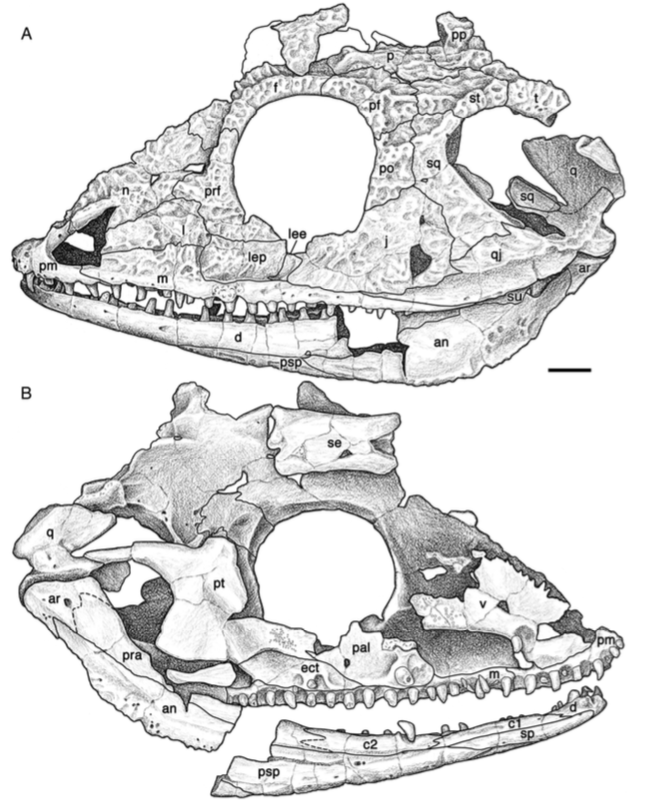
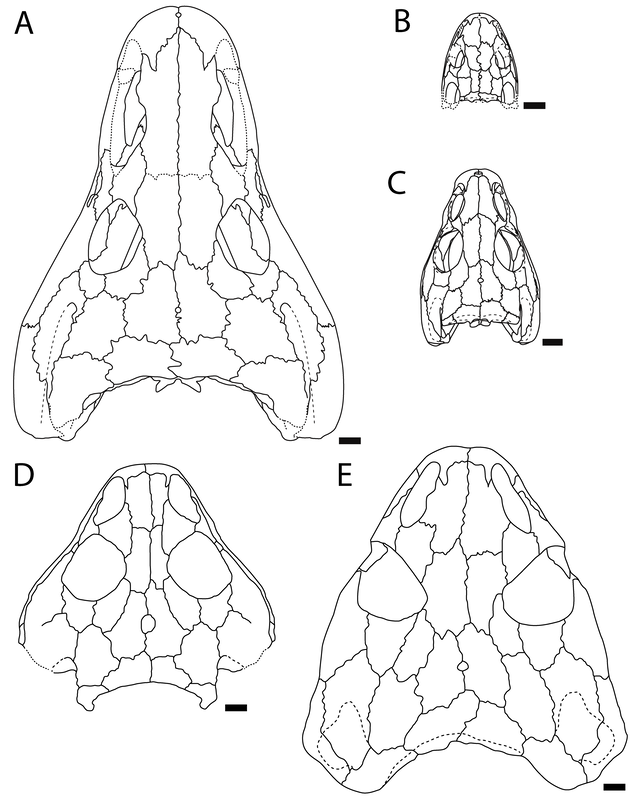
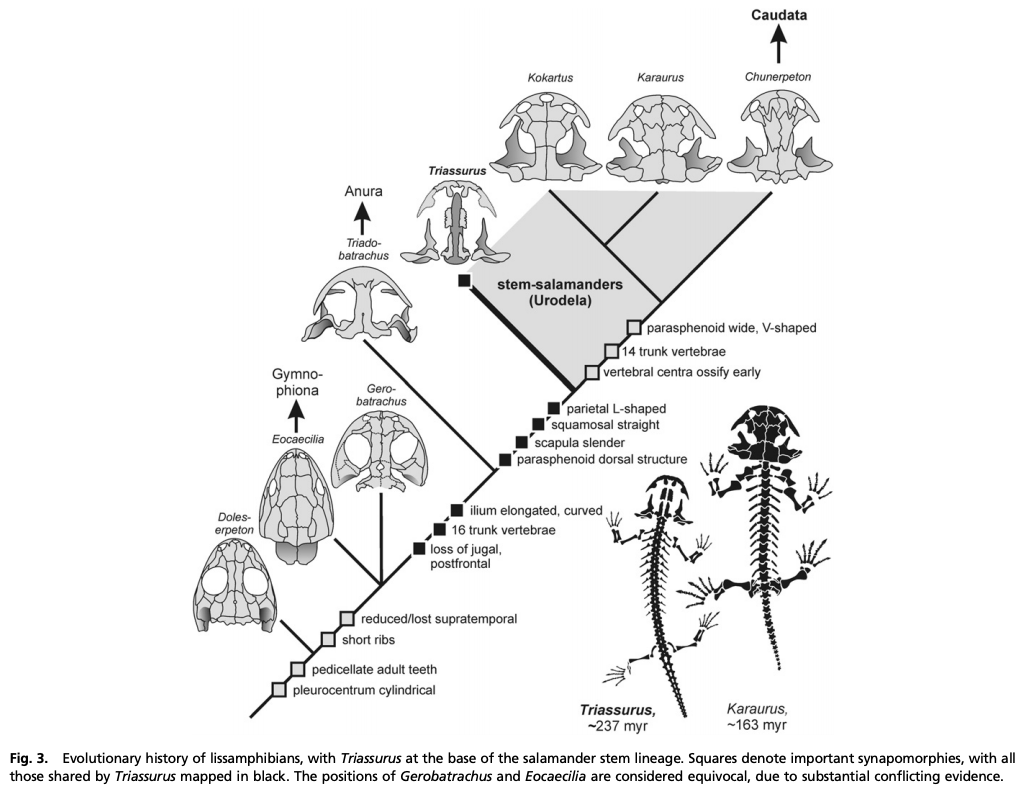 The second study was a broader review of the fossil record of caecilians by Santos et al. (Biol J Linn Soc); the second author (Michel Laurin) is a longstanding proponent of a single origin of lissamphibians from lepospondyls. This paper repeatedly calls out various lines of argument used by Pardo et al. and even has a discussion section titled "Chinlestegophis: a true gymnophionomorphan?" In fact, they go so far as to say that every identified synapomorphy is questionable, and other shared features are plesiomorphies found in other temnospondyls. The issue of whether the element at the front of the eye is a lacrimal or a LEP is just one of those questioned features. No new phylogenetic analysis here, but a great summary of the available information on fossil caecilians (it is very poor) like the figure below on the left. In case you're wondering why the record is so bad, burrowing animals by virtue of where they live are not nearly as likely to end up near the types of environments that most frequently preserve in the fossil record, like ponds or floodplains. Their small size, like that of other lissamphibians, is another issue that is generally less favourable for preservation.
The dissorophid dynastyThe temnospondyls long considered to be closely related to lissamphibians in one form or another, dissorophoids, continued to see major research interest, even outside of the context of lissamphibian origins. 2020 mostly brought re-descriptions, though we got a few other types of studies thrown in there as well.
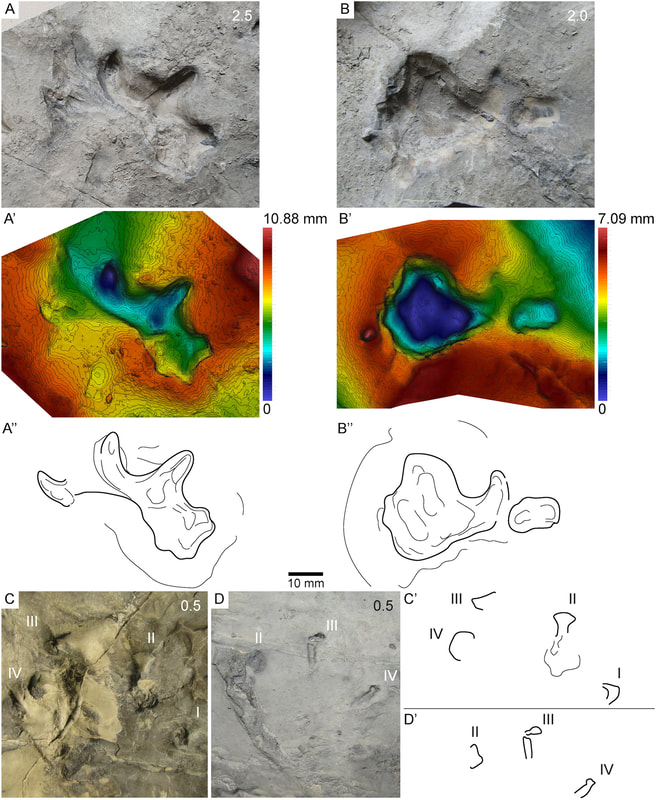
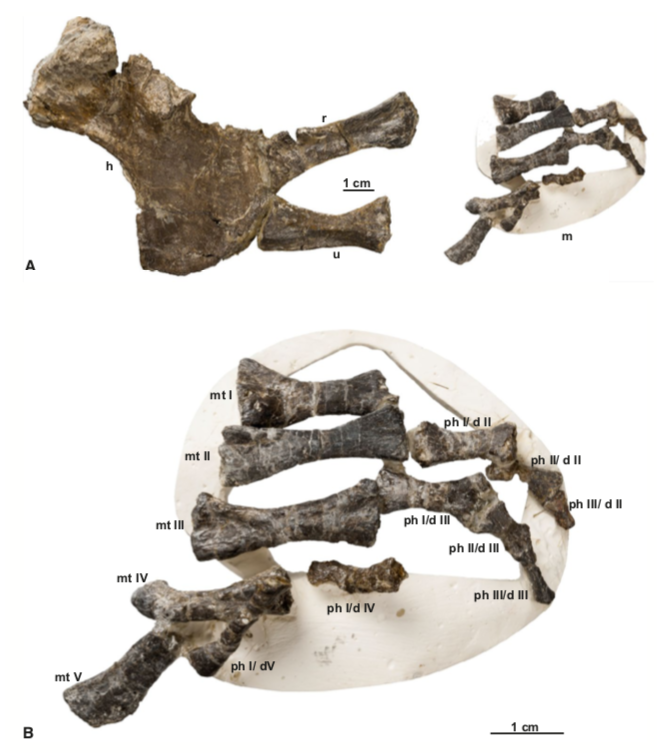
One important redescription that we got in 2020 was of the holotype of "Dissorophus" angusta, a species named by Bob Carroll in 1964. Basically everybody has recognized for decades that it is definitely not Dissorophus, lacking the large first osteoderm and having narrow osteoderms throughout the body, but it is only now, 56 years later, that its status is finally resolved as a new genus, Diploseira, by Dilkes (J Vert Paleontol). The only known specimen is mostly the postcranial skeleton, which makes it hard to compare with many other dissorophids, but it definitely has distinctive features. David's usual impeccable attention to detail and illustrations really flush out every little nuance of the anatomy. 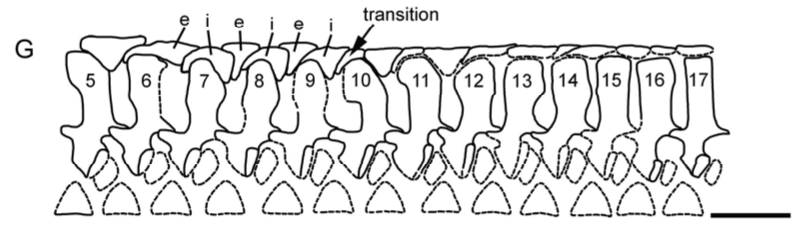 This taxon is quite interesting in preserving a transitional series of osteoderms; there are two series at the front and only one at the back. As far as we know, this is the only dissorophid with this condition - other species either have a single continuous series or two continuous series (external and internal). Much of the osteoderm anatomy (though not the width) is shared with dissorophines (Broiliellus and Dissorophus), and Dilkes' phylogenetic analysis indeed recovered Diploseira within that group. This really mucks up how we use qualitative patterns of osteoderms to make taxonomic frameworks, continuing to highlight the issues with single-feature phenetic taxonomy (essentially arbitrary emphasis of certain observational data).
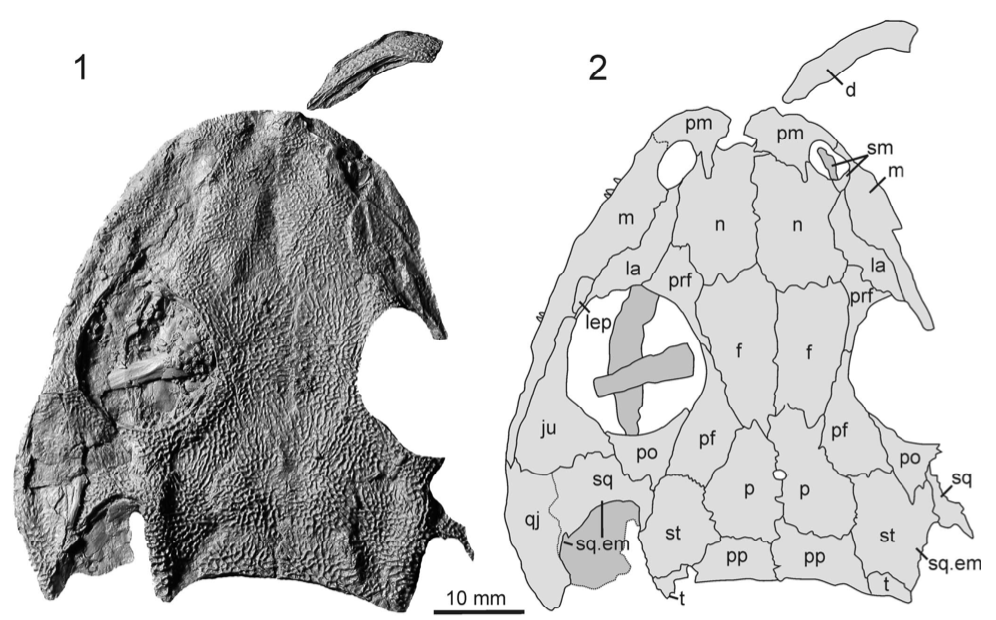
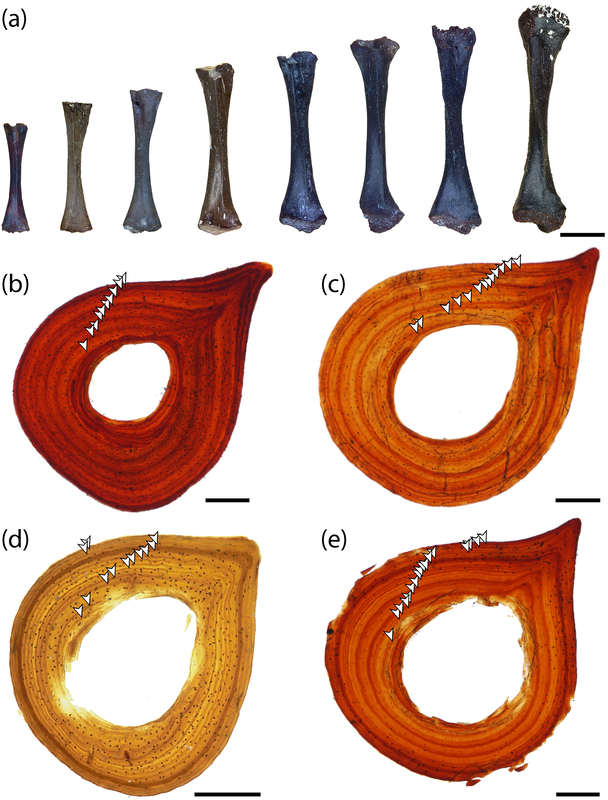
A slice of lifeIn addition to my Doleserpeton paper, there were a few other studies that sliced and diced up some temno bones. The first one was technically published online last year (in print this year), and I very briefly mentioned it in my year-end summary last year, but it's worth discussing here in greater detail. Mukherjee et al. (Papers Palaeontol) describe the histology of a few Middle Triassic capitosaurs from India, which is a follow-up to another paper led by Muhkerjee a decade ago on a more preliminary sample of Indian temnos. Despite the material of the two taxa (Cherninia and Paracyclotosaurus) coming from the same small locality, they show distinctive differences in their bone histology. Cherninia shows a lot of what temnospondyl workers term 'incipient fibrolamellar bone,' which generally meets the criteria of fibrolamellar bone in amniotes. Note that contrary to some oversimplifications, fibrolamellar bone is not an unequivocal hallmark of endothermy - it only indicates rapid growth, which can be accomplished in ectotherms living in harsh conditions that require fast growth (for example, taxa with larval forms that need to get out of water before it dries up).
Comparison of histology of the humerus of Cherninia (on the left) and Paracyclotosaurus (on the right). In Cherninia, this fibrolamellar bone tissue is found in sub-adults too (it would be more likely to be found in juveniles that are still growing), but it is not found in any growth stage of Paracyclotosaurus, which also lacks the woven-fibered bone of the smallest and most rapidly growing individuals of Cherninia. So Cherninia = fast grower, Paracyclotosaurus = slow grower. The histological / microanatomical differences can be correlated with distinct differences in the proportion of limb features and the torsion of the humerus and femur, suggesting biomechanical differences in their locomotion. The authors proposed that Cherninia was a classic obligately aquatic taxon, like what we think of most stereospondyls, whereas Paracyclotosaurus was more capable of moving around on land, not like what we typically think of large stereospondyls. It's debatable in my opinion whether the latter "spent a considerable amount of time on land," but it definitely seems likely that being able to at least move between ponds would have been advantageous, and it suggests that there may be a lot of cryptic information to be derived from the histology and microanatomy that is obscured by more conserved external anatomy.
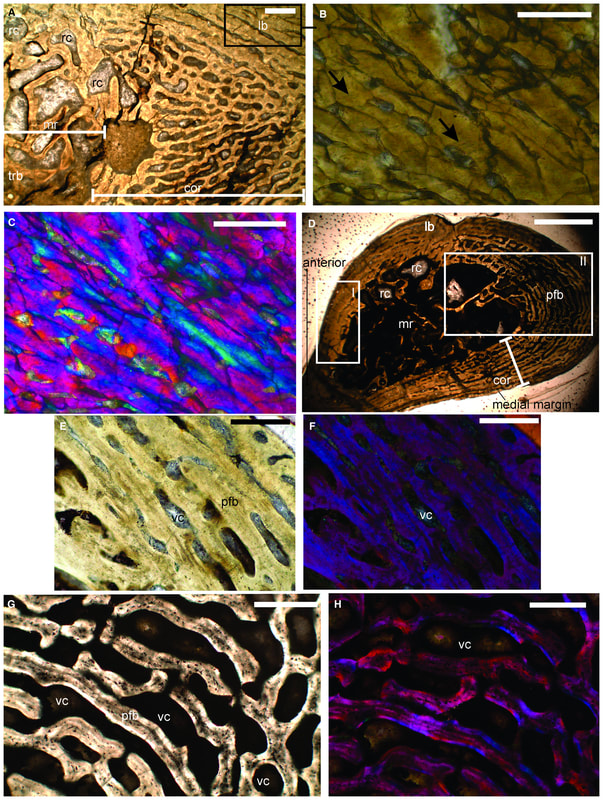
Comparison of sections of the humerus of Panthasaurus (on the left) with the femur, ulna, and tibia (on the right; A-H = femur; I-J = ulna; K-M = tibia). 2020 was the first year that I didn't have any new metoposaurid papers published (there's stuff in the pipeline, don't worry), but the metoposaurid histology train rolls on. Teschner et al. (PeerJ) reported histology and microanatomy of the Indian metoposaurid, Panthasaurus. This taxon shows pretty normal features for obligately aquatic stereospondyls, like parallel-fibered bone with remodelling that's accompanied by lamellar bone tissue (these reflect slower rates of growth), a fairly poorly vascularized cortex, and distinct growth marks (but not LAGs). This study is really nice because it adds to the growing body of literature helping us to understand how closely related taxa (within the same family) might differ in ecology or response to local climate, things we might not be able to nuance out from the external anatomy alone. Years ago, there was a nice comparative study by Dorota Konietzko-Meier (who's on this paper) and Nicole Klein comparing the signals in Dutuitosaurus from Morocco and Metoposaurus from Poland, which showed that the Polish paleoenvironment was milder and led to slow-downs or short stagnations but not long cessations in growth. Panthasaurus shows a similar signal to Metoposaurus (growth zones and annuli but no LAGs), indicating the Indian climate was also not too harsh. The authors also sampled a bunch of different elements, which is a key step forward to expanding the utility of non-limbs for histological studies by providing a reference point that isn't a humerus or a femur; if you wonder why nobody has done any of this work on North American taxa, it's because there are very few limb bones that are basically off-limits to destructive sampling right now (a number of people other than me have looked around, and this is why I keep cutting intercentra instead). Finally, Uliakhin et al. (Paleontol J) reported what is probably the highest age estimation for any Paleozoic vertebrate and definitely the highest one for any temnospondyl - a whopping 57 years! This was reported from the late Permian dvinosaur Dvinosaurus campbelli from Russia; dvinosaurs are one of the few uncontroversially fully aquatic Paleozoic temnospondyls, and there remains fringe speculation that one of the stereospondyl clades might actually be tied to this group. We see a lot of the same features that typify stereospondyls (e.g., parallel-fibered bone, lots of secondary remodeling from the medullary cavity, calcified cartilage), which is, at minimum, a reflection of what happens when you're neotenic and end up permanently living in the water. 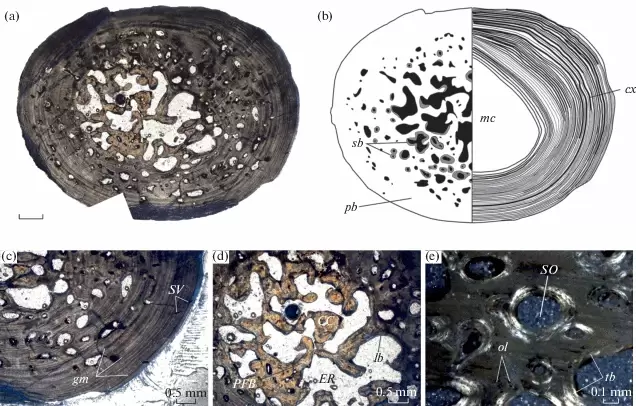 How about the 57 years - is that out of bounds? We know that slow-growing animals tend to live longer (why grow slow if you'll die in a year), and some modern amphibians top 60 years (and it's not one of those weird one-offs that far exceeds the norm for the species). Like the authors point out, neotenic individuals tend to live longer than metamorphosing ones. Of course, this assumes that the interpretation of the LAGs is correct; could it be double LAGs (two cessations in growth per year) for example (thus halving the estimated age)? Hard to say. The giveaway for double LAGs is their spacing - two closely spaced lines with a gap smaller than that from the line on either side. That doesn't mean you couldn't get double LAGs that are spatially indistinguishable from normal LAGs if the process that formed them was timed differently. For example, the way that we define seasons usually sets the start of winter and the start of summer as exactly 6 months apart. But as most people experience them, the "peak conditions" of each season aren't on Day 1, and the peaks may not be 6 months apart. Growing up in SoCal, I would say the hottest month of the year can be as late as September, and having lived in Canada, February or March can be the coldest. That asymmetry is likely what produces the characteristic spacing of double LAGs. But of course there's variation - who really knows the finer nuances of the climate in Russia over 250 million years ago, after all. So it's possible that climate peaks really could have been nearly 6 months apart and thus produce a pattern resembling single LAGs when the animal actually stopped growing twice a year. But we can't really prove that either. The best way to interpret this would be to cautiously accept the author's interpretation (single LAGs = 57 years) but not to make too much of it. Comparing maximum ages in extinct taxa is hard because how do you know that you got one that died around its maximum age? A lot of animals (especially those lower on the food chain) frequently die well before their maximum age, often because they're eaten, so the fact that other temnospondyls rarely exceed estimates of 15 years only says something about the sample, not necessarily the species as a whole. Piling up
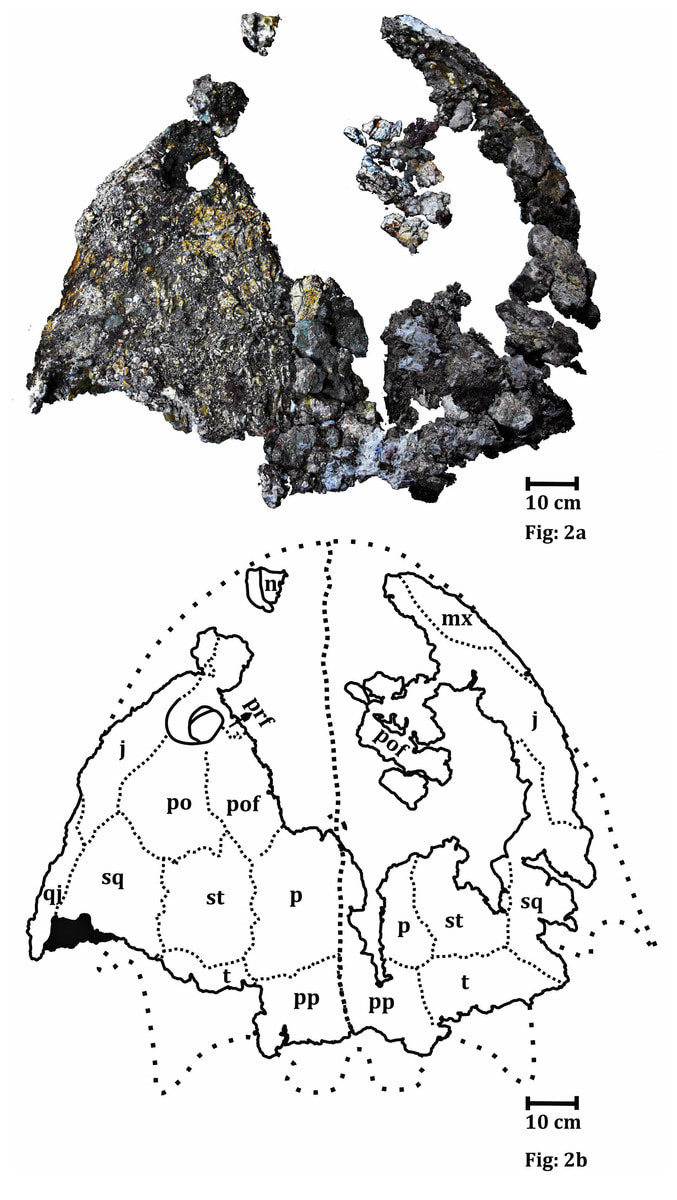
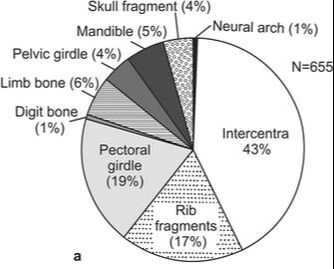
Based on a slew of taphonomic analyses, there appears to be disparate pre-burial conditions between the different tetrapods found at the site that explains the differences in their representation (the complex graph on the top left). There are mixed age classes for the metoposaurids, whereas the phytosaurs and rhynchosaurs are skewed towards either juveniles or adults. Obviously some of this has to do with which animals lived in the immediate environment versus which might have been carried in and the cause of death. This is not a mass death assemblage like that of Dutuitosaurus or the Lamy quarry insofar as it appears to be time-averaged for metoposaurids (but not rhynchosaurs, which may have mostly gotten swept in by a single event), and instead seems to be accumulation of their remains through periodic flooding and a build-up of remains in low-energy spots; the model is on the top right. The authors went so far as to argue that the metoposaurid remains might have been left out on the banks, exposed for years and constantly weathering, which might account for the total lack of complete skulls. Really great prospects based on the sheer volume of material that's come out of the site - hope to see more in the future! That's gotta hurtWith advances in technology and application of modern methods to paleontological specimens, paleopathology (studying diseases in fossils) has come a long way - we're seeing more and more reports of specific diagnoses and maladies beyond "that is clearly not right." Novikov et al. (Paleontol J) describe a bone lesion in the Early Triassic trematosaur Benthosuchus. This is one of the most common Early Triassic tetrapods found around the world (but mostly in Russia in this case), and the authors report a bone lesion on the lower jaw, which would be immediately identifiable to even non-scientists as a very odd-looking round protrusion from the side. 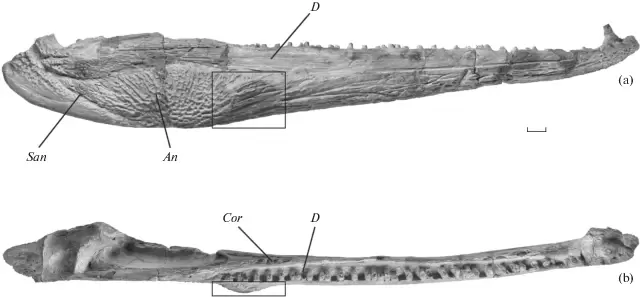 Based on features such as denser structure compared to surrounding bone, localized presence of a smooth feature, and no apparent connection to the teeth, the authors propose that this represents a non-odontogenic osteoma (non-tooth-related tumour). Because they couldn't do histology on the jaw, their results are based only on the external examination and the CT data (not the sharpest), so there are a few other possibilities (like a cyst or specifically a bone cancer). This remains the oldest example of a tumour forming in a tetrapod, although other reports like my friend Yara Haridy's 240-million-year-old turtle tumour are not that much later. Welcome to the club
A few other temnospondyl adjacent studies:
And the recent special issue on Karoo biozonation: https://pubs.geoscienceworld.org/sajg/issue/123/2 (note there are a few included taxa that are probably junior synonyms in some articles). In closingAll things considered, an excellent year for temnospondyl research! I reviewed a bunch of these papers as well, which is always neat to see what people are working on ahead of press. It's been a few months since I had anything come out (a coincidental pile-up of my 2019 productivity in the first half of this year), but I've got a few things working their way through the pipeline and will hopefully be getting back to some semblance of regular blogging in the new year! Thanks for reading, and best wishes for the new year! References
|
About the blogA blog on all things temnospondyl written by someone who spends too much time thinking about them. Covers all aspects of temnospondyl paleobiology and ongoing research (not just mine). Categories
All
Archives
January 2024
|







 RSS Feed
RSS Feed
