|
If you look at a temnospondyl skull, it is decidedly not smooth like our own. Instead, it may be covered by any (or all) of the following: pits, grooves, ridges, tubercles, or nodules. Don't confuse this for pebbly or bumpy skin texture like in some modern reptiles that are formed by separate scales or osteoderms that sit on the skull (or other parts of the body) - these bumps develop as part of the skull! This week's blog post is covering dermal ornamentation, naturally with a focus on temnospondyls. A lot of this week's blog is derived / inspired by work by Florian Witzmann at the Museum für Naturkunde in Berlin who has done lots of very cool work on temnospondyls in general but also with respect to their bones.  Mounted skeleton of the Late Triassic temnospondyl Koskinonodon perfectus from Texas (labeled as Buettneria perfecta) at the American Museum of Natural History in New York. Ornamentation is developed on the top of the skull and the side of the jaw. Photo: Bryan Gee  Photograph of the Gila monster (a lizard from Mexico), which has a pebbly texture as a result of scales and osteoderms rather than well-developed ornamentation. Distributed on Flickr by u/OzinOH under a Attribution-NonCommercial 2.0 Generic (CC BY-NC 2.0) license. Who has it? Temnospondyls are well-known for having heavily ornamented skulls, especially when they occur in deposits with other tetrapods that are largely or entirely un-ornamented (e.g., Late Triassic deposits) This can make it pretty easy to tell whether a temnospondyl was around and who it was, even just from small fragments. However, they are not the first or the last tetrapods to be ornamented - many Paleozoic tetrapods (e.g., stem tetrapods, seymouriamorphs), including some amniotes (e.g., captorhinids, crocs [both living and extinct]), are ornamented to various degrees. Below are some examples of ornamented non-temnospondyls. Where is it? In temnospondyls, the most apparent ornamentation is found across the skull, in which most to all of the elements are ornamented on their dorsal (top) or lateral (external side) surfaces. This is the most common place for ornamentation in non-temnos as well. Temnospondyls also have ornamentation on parts of the jaw (again, common for early tetrapods), particularly along the ventral (bottom) margin and toward the back on the lateral (outer) surface. Lastly, many (but hardly all) temnospondyls have ornamentation covering the ventral (bottom) surface of the main plate-like pectoral (shoulder) elements, specifically the interclavicle (a bone not found in mammals) and the clavicle (the collarbone). These pectoral bones face downward in quadrapeds (4-legged locomotion) and towards the front in bipeds (2-legged locomotion). A few temnospondyls have osteoderms (bony armour plates), which also ornamented (osteoderms are typically ornamented in non-temnos). Ornamentation can differ within the same animal at virtually every scale - between skeletal regions (skull vs. osteoderms), within regions (snout vs. back of skull), and within element (e.g., parietals). What's it for? Because ornamentation is so ubiquitous, it has been widely (although not extensively or conclusively) speculated on. Below are some of the main ideas behind ornamentation:
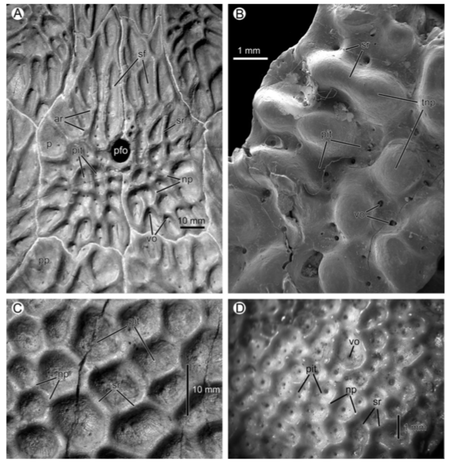
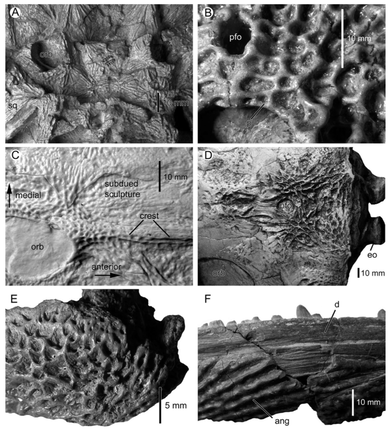
Some of these ideas hold more water than others, and a number of them are related to the widespread consensus that a complex and well-developed vascular network was integrated with the ornamentation. Evidence for the vasculature is evidenced through small foramina that are found throughout the ornamentation (see figures below).
Cutaneous respiration Breathing through the skin, or cutaneous respiration, is a common physiological attribute of modern amphibians, hence why they often need to keep their skin wet and usually do not do too hot in deserts and other arid climates. Other animals (including humans) are capable of cutaneous respiration, but this typically is responsible for a very small amount of our total respiration, whereas it is pretty important for amphibians. The most speciose group of salamanders (plethodontids) are called lungless salamanders because they actually lack lungs and more or less rely entirely on cutaneous respiration (e.g., below on left)! Bystrow (1947) was one of the first proponents of the idea that ornamentation of various tetrapods was primarily for cutaneous respiration. There are a few problems with this however...
Buffer against gas buildup This is a relatively novel idea put forward by Janis et al. (2012) that ornamented dermal bone is more effective at buffering acidosis (buildup of acids in the blood), primarily of carbon dioxide (turns to carbonic acid in the blood) and lactic acid. The idea is based on modern vertebrates and largely (well-supported) conjecture about the evolution of early tetrapods and the associated physiological challenges. As pointed out by the authors, fish (living in a lower-oxygen environment) need more ventilation to pull in oxygen through their gills and thus get rid of carbon dioxide much faster due to high ventilation rates. Conversely, terrestrial animals (living in a higher-oxygen environment) ventilate at lower rates and thus accumulate more carbon dioxide. This is a particular problem for larger animals. Various authors have speculated on how early tetrapods eliminated carbon dioxide (e.g., Packard, 1976). Although some of the more aquatic early tetrapods may have retained external gills and been able to dump CO2 by returning to the water, this would not work for a fully terrestrial animal. Costal (rib-driven) respiration like that seen in reptiles also seems unlikely because a mobile ribcage had not evolved. They also probably lacked other mechanisms seen in amniotes (e.g., buffering with the kidneys); modern amphibians are not very efficient at compensating for acidosis. In modern tetrapods, mobilization of calcium deposits from bone tissues can be used to buffer acidosis. In general, predicted patterns are supported by the fossil record. Terrestrial tetrapods have more pronounced ornamentation on the whole compared to aquatic forms, except for ones that are comparably small to modern amphibians and that probably could buffer through increased gas exchange through un-scaled skin. Ornamentation is also rare in early amniotes, indicating that costal respiration may have evolved by this time. A few problems:
Verdict: Appealing but difficult to test.
Improved stress distribution Coldiron (1974) was the first major proponent of the hypothesis that developed ornamentation helped to better distribute stresses, particularly during feeding. This is an attractive idea with respect to the skull, but it does not account for ornamentation on other elements (particularly the postcranial ones) that are not exposed to similar stresses. Additionally, the directionality of ornamentation with a particular orientation (e.g., ridges) does not align with predicted stresses but rather with the development of the vascular network. A more recent study by Rinehart and Lucas (2013) used a more unusual analytical method to survey two Triassic temnospondyls (Eocyclotosaurus, Koskinonodon): beam mechanics. Modeling two main types of ornamentation, reticulate (honeycomb pitting), and ridge-and-groove, they showed that these types of sculpturing provide different resistances to various stresses and proposed that distribution may relate to muscle attachments (extending to the pectoral elements). This is an interesting finding based on an entirely different approach (P.S. I know nothing about beam mechanics), but it assumes more about temnospondyl musculature than is presently known and certainly requires more testing and taxon sampling. Verdict: Unlikely but needs more testing
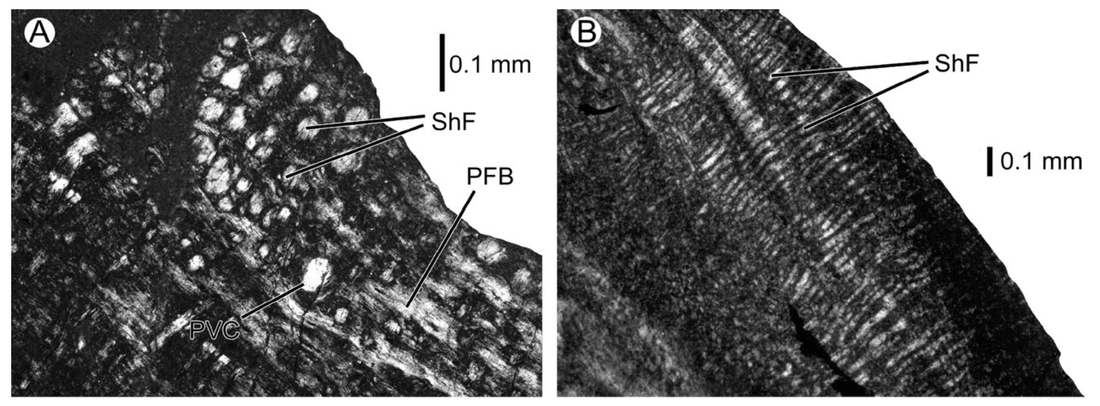 Thin sections of sculptural ridges in temnospondyls showing Sharpey’s fibres (ShF) penetrating the bone. A, Chenoprosopus milleri; B, Plagiosternum granulosum. Figure from Witzmann et al. (2010). Indicator of metamorphosis Boy & Sues (2000) found a correlation with development of polygonal ornamentation with remodelling of the hyobranchial apparatus, which occurs during metamorphosis. This sort of makes sense - many parts of the skeleton change in various ways during metamorphosis in modern amphibians, and this may well have been true for temnospondyls. Metamorphosis has been documented in a number of temnospondyls (e.g., Schoch, 2002, 2009). A few problems...
Verdict: Possible but only for particular clades for which metamorphosis is documented.
Is temnospondyl ornamentation diagnostic? Yes and no. Some temnospondyls have pretty weird ornamentation. For example, plagiosaurids (e.g., Plagiosuchus pustuliferus, see higher up the slideshow on temno ornamentation) have a pretty recognizable pustulated texture on their elements that is formed by densely packed tubercles (pustules are supposed to have pus in them), in contrast to the more typical pitting and grooving of other temnospondyls. Isolated, ornamented temno chunks can still be diagnostic or at least informative as well, either for guessing at what genus or species the fragment belongs to or what element(s) it comprises. For example, metoposaurid ornamentation is mostly honeycomb patterning made of semi-circular pits, but elongate grooves are found on the parietals. In general though, it can take a lot of observations and practice in the field to be able to identify fragments on the fly, and good luck if you get a random frag with just a few circular pits on it. Is temnospondyl ornamentation difficult to prepare? Sometimes accretion of hard minerals to ornamented pieces makes it hard to prepare, especially with ridges and tubercles (3D projections). This is why a lot of the specimens collected in the 19th and early 20th centuries don't have well-preserved ornamentation (it can also be weathered to be flatter when exposed). Pitting (3D depression) is usually a little easier to prepare because sometimes the infilling just pops out. I've actually exposed entire skulls in which the impression was preserved as a single piece on the rock that was lying on top of it. Conversely, sometimes you spend weeks just scraping the pits and grooves to get out residual pieces of matrix from when you put too much glue on the specimen...
Closing thoughts Like most things in paleontology, it isn't easy to conclusively say what led to the evolution of ornamentation in early tetrapods, and there are probably a multiplicity of environmental and ecological factors at play in addition to phylogenetic constraints that produce the diverse array of ornamentation that we observe in the fossil record. Up next week: TBD, some kind of profile on a particular temno group Refs
Edit (1/22): minor corrections based on comments by D. Marjanović.
David Marjanović
1/17/2019 11:45:43 am
"In modern tetrapods, mobilization of calcium carbonates from bone tissues can be used to buffer acidosis." Comments are closed.
|
About the blogA blog on all things temnospondyl written by someone who spends too much time thinking about them. Covers all aspects of temnospondyl paleobiology and ongoing research (not just mine). Categories
All
Archives
January 2024
|







 RSS Feed
RSS Feed
