|
Busy week, so just a short post this week about some bones in the eye (yes, they actually are in the eye); it's not entirely random - Lars Schmitz, my former anatomy professor, is coming to UTM to give a talk on the evolutionary signals in vertebrate eyes (an unsurprising topic selection if you know him). 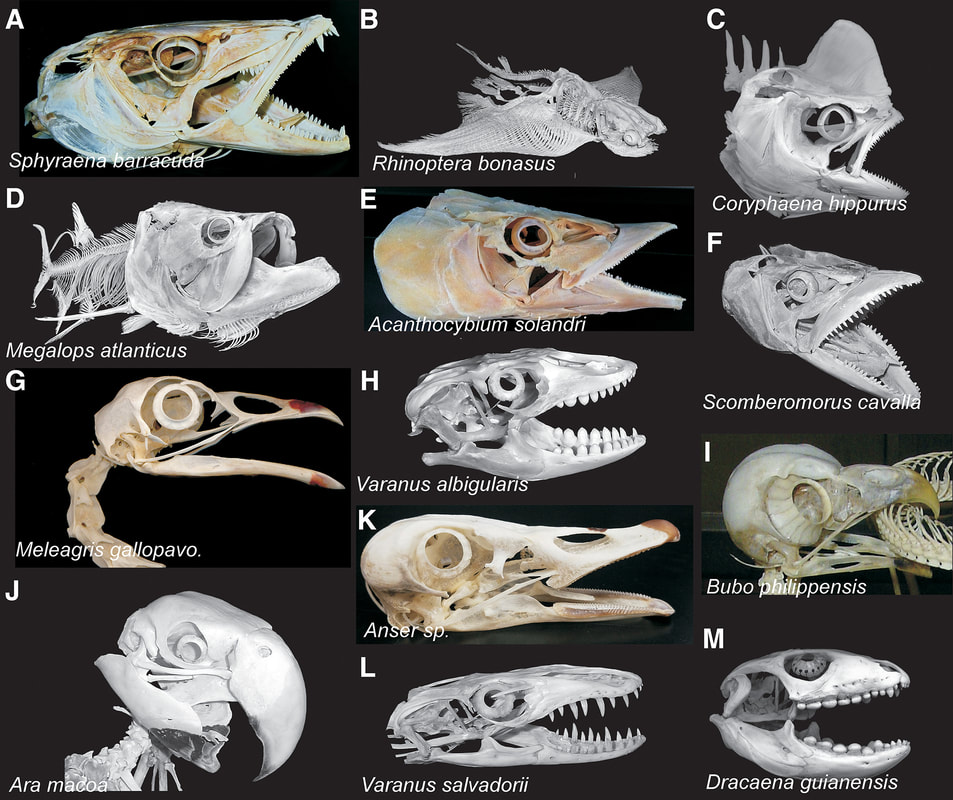 Examples of scleral rings in vertebrates. Common names in alphabetical order of images: great barracuda, cownose ray, mahi mahi, tarpon, wahoo, king mackerel, wild turkey, rock monitor lizard, eagle-owl, scarlet macaw, goose, crocodile monitor lizard, caiman lizard (figure from Franz-Odendaal, 2018)
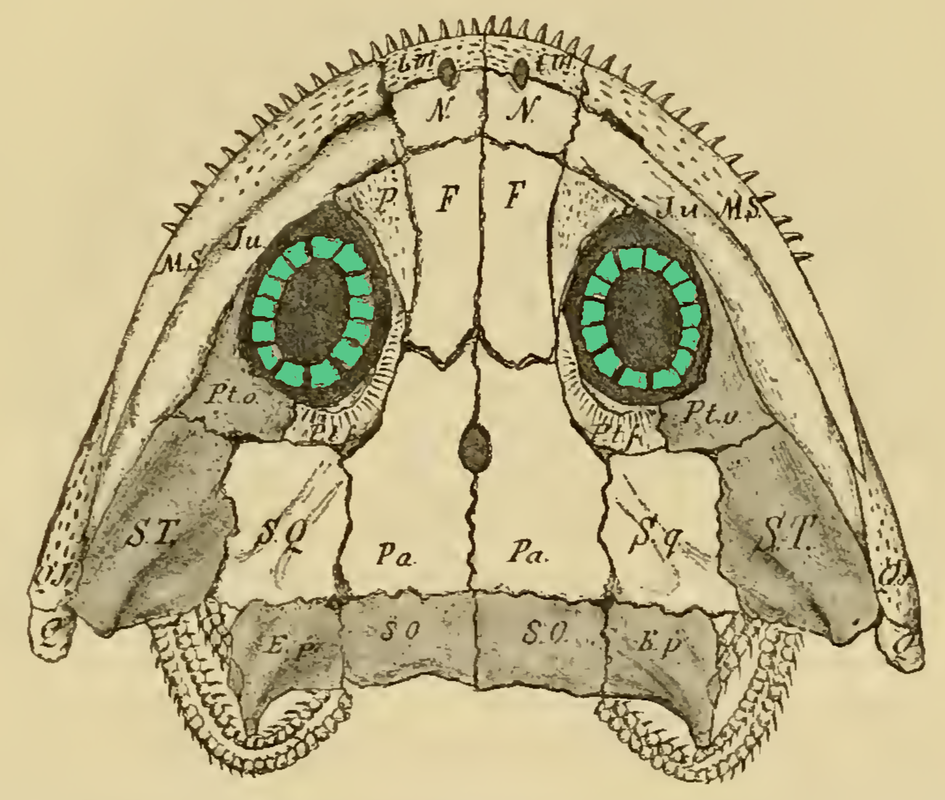
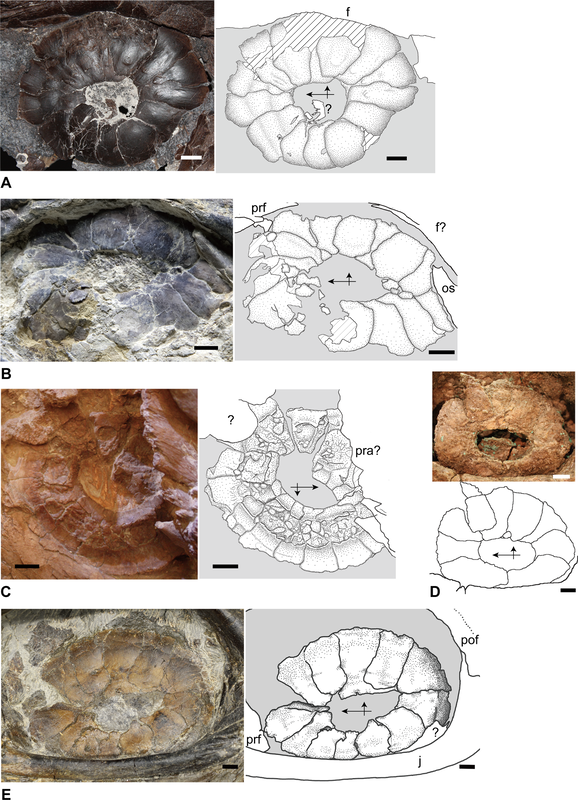
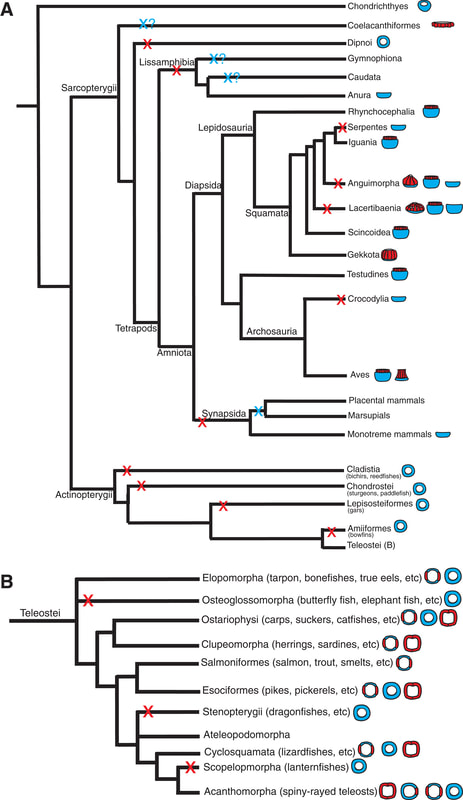
How do you differentiate a scleral ring plate from a palpebral bone (or fragments of it)? Obviously when either is articulated or fully complete, it's a little easier to differentiate them. When they're not, palpebral bones are often ornamented as well, like the skull roof, whereas scleral rings are smooth. The palpebral bone also tends to break into pieces of various sizes, whereas the plates just disarticulate but are otherwise of about the same size. So what do we actually know about these ossifications' function in temnospondyls? There isn't very much research into the eye ossifications in temnospondyls because they occur fairly rarely and are often broken or disarticulated. Because the scleral ring comprises many plates that disarticulate after death, and the palpebral bone seems to break readily into many pieces, loss during preservation is highly likely, which compromises the use of their presence / absence in phylogenies, similar to other small and mostly unattached bones. The palpebral bone probably acted in a similar fashion to those in living vertebrates where it mainly supports the eyelid. The function of temnospondyl scleral rings are more difficult to infer. They occur in both large and small, aquatic and terrestrial temnospondyls, so like many other aspects of their anatomy, they may serve relatively limited functions and be inherited through phylogeny. Although it's appealing to try to draw some sexy ecological inference from what appear to be relatively large rings and relatively large orbits in temnospondyls, different selection pressures affect and maintain scleral rings (i.e. there is no simple explanation), so aquatic versus terrestrial animals probably evolved scleral rings with different primary functionalities, sometimes for improving vision, sometimes for reinforcing the eye, muscle attachment for accommodating the lens, etc. Presence / absence and number of sclerotic plates in other groups of vertebrates is sometimes correlated with differing ecologies; deep-sea and relatively inactive fish, for example, either lack scleral ossicles altogether or reduce the number of these ossicles (e.g., Franz-Odendaal, 2008). The relative size of the scleral ring to the eye has also been used to infer activity patterns of various terrestrial animals (e.g., Schmitz & Motani, 2011, see below figure). Unfortunately, sclerotic ossicles are too uncommon (and rarely articulated) in the temnospondyl record to make much of them, and modern amphibians have lost them (but do occasionally have other ossified structures), so there isn't a good living homologue (or analogue). They may just as well as served a function for maintaining the structure of the eye in temnospondyls. 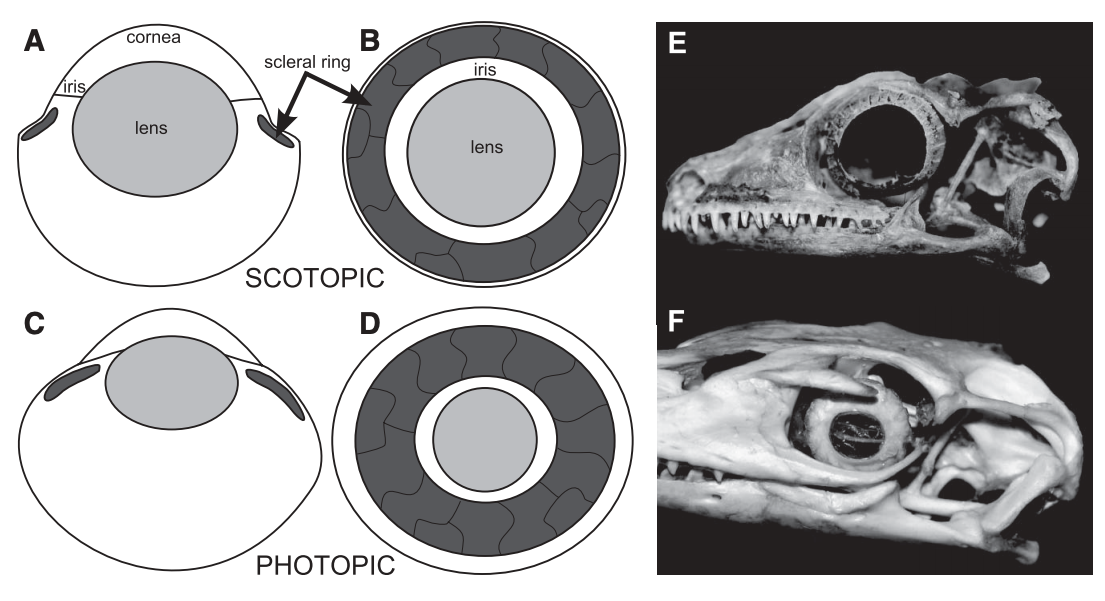 Comparisons of the scleral ring in scotopic (nocturnal) animals, with the gecko Rhacodactylus (E) as an example, and photopic (diurnal) animals, with the monitor lizard Varanus as an example, figure from Schmitz & Motani (2011). There's been a ton of work looking at the sclerotic ring in particular as a proxy for inferring activity patterns (diurnal, nocturnal, etc.) in other tetrapods! If you're looking for some good (and short) examples:
Refs
David Marjanović
3/7/2019 11:42:18 am
It looks like the sclerotic ring is lost most often in nocturnal clades like mammals and crocodiles. This fits your last figure, of course. Comments are closed.
|
About the blogA blog on all things temnospondyl written by someone who spends too much time thinking about them. Covers all aspects of temnospondyl paleobiology and ongoing research (not just mine). Categories
All
Archives
January 2024
|

 RSS Feed
RSS Feed
