|
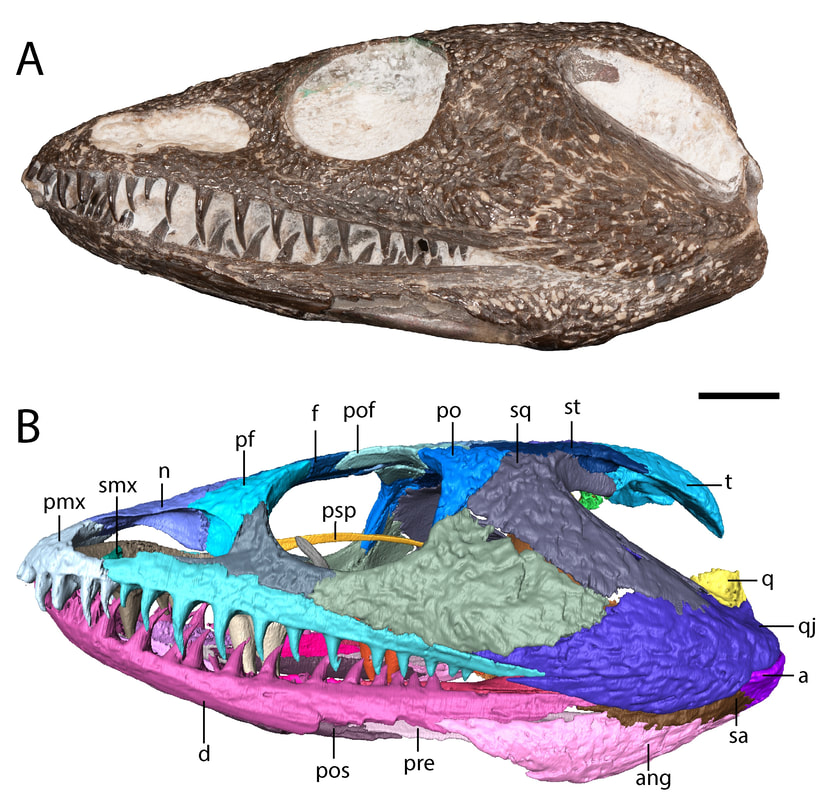
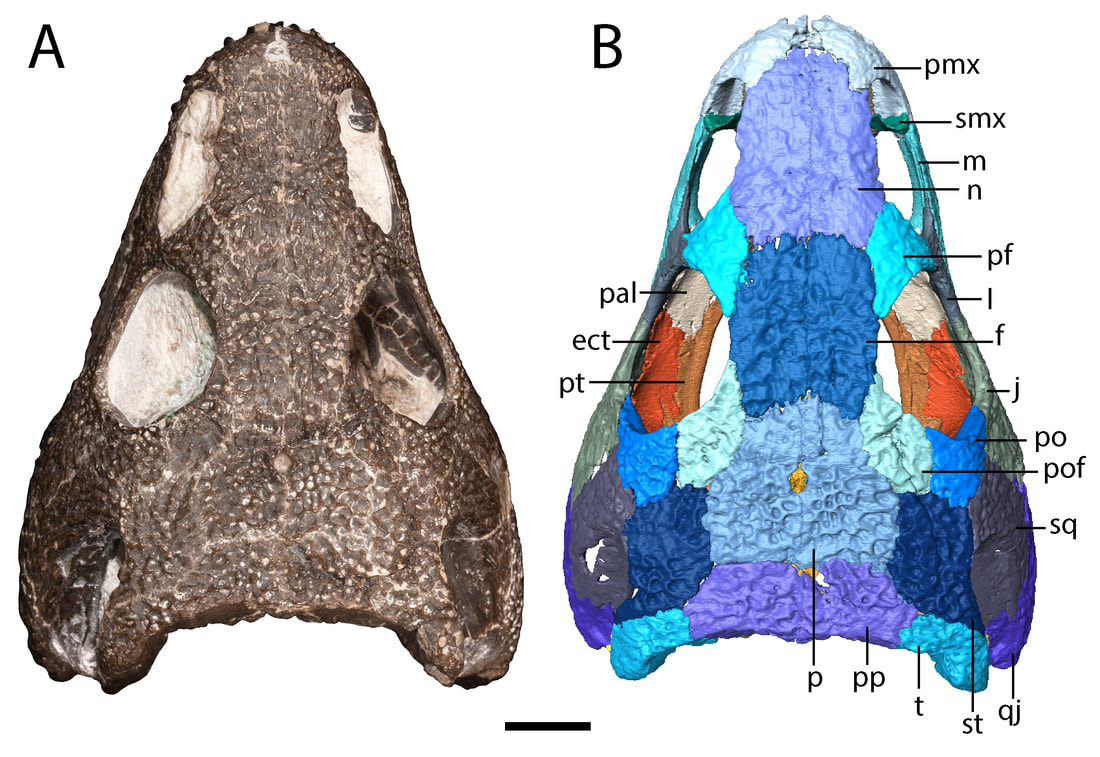
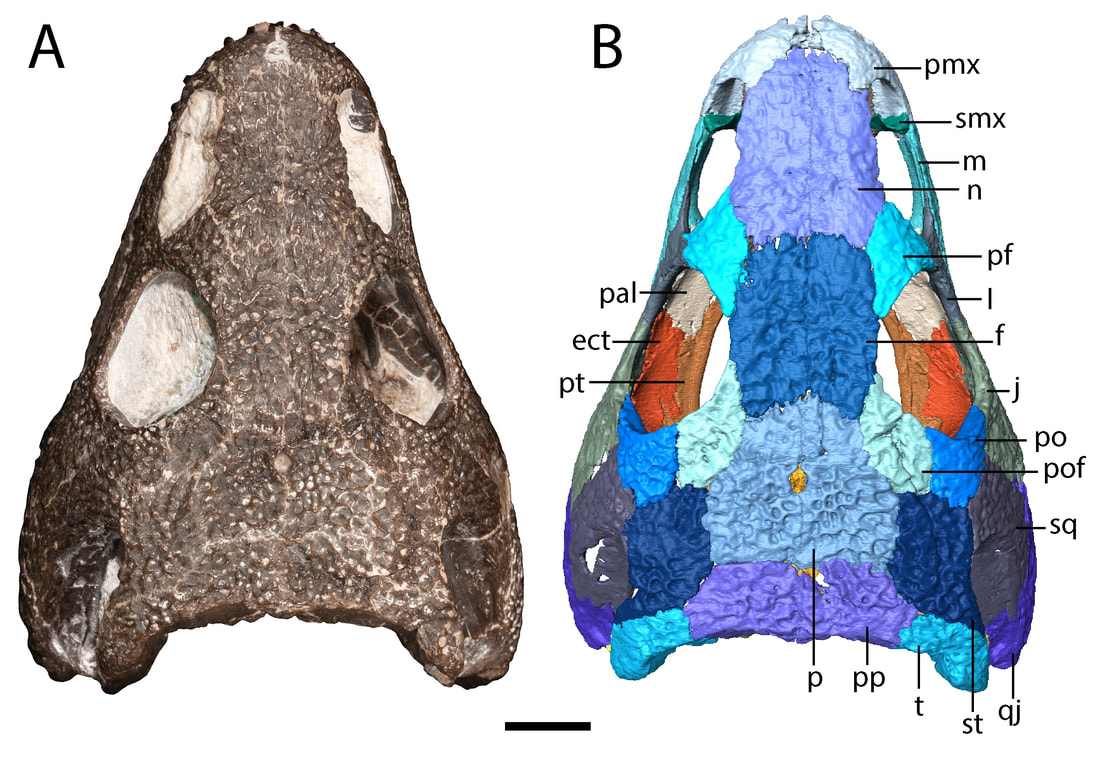
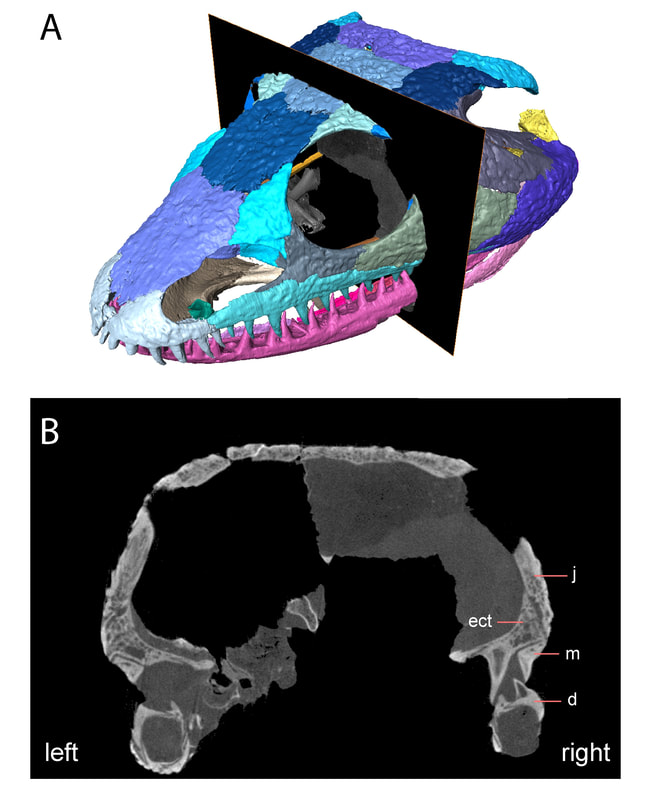
Title: A juvenile specimen of the trematopid Acheloma from Richards Spur, Oklahoma and challenges of trematopid ontogeny. Authors: B.M. Gee, J.J. Bevitt, R.R. Reisz Journal: Frontiers in Earth Science, v. 7, p. 38 Link to paper - this is an open access paper than anyone can read! 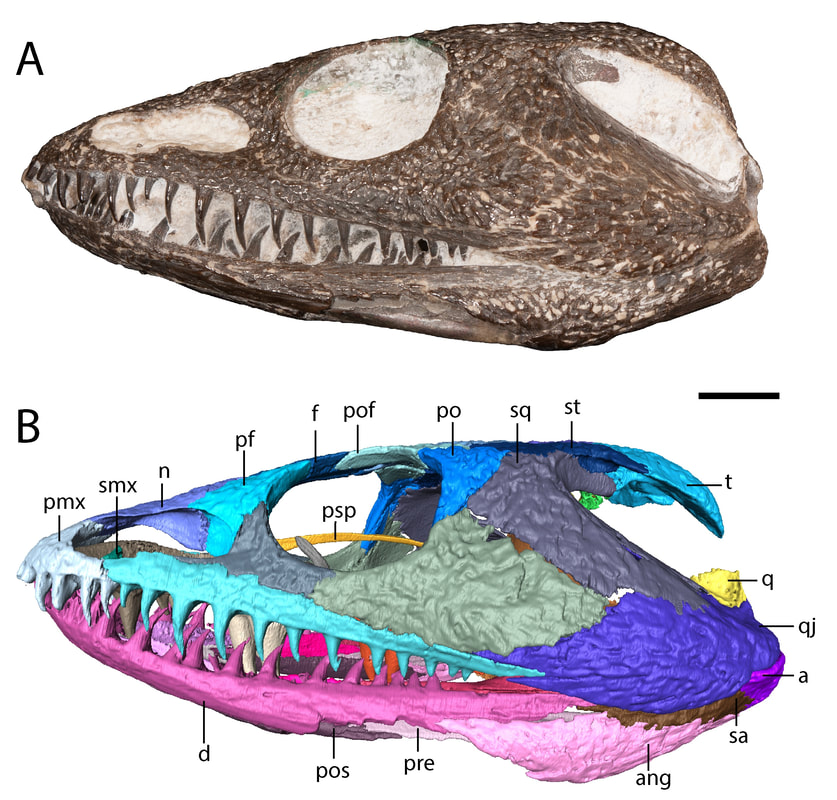 Photograph and digital segmentation of the small skull of cf. Acheloma that we describe here (Figure 2 in the paper); scale bar = 1 cm. Colorized surfaces follow my previously used CT colour palette. Abstract (from paper) Trematopids are a clade of terrestrial dissorophoid temnospondyls documented primarily from terrestrial Permo-Carboniferous environments in North America and Europe. Here we describe the complete skull and articulated mandibles of a diminutive trematopid specimen (OMNH 79318) from the Early Permian karst deposits near Richards Spur, Oklahoma. Based on aspects of the neurocranium (e.g., unossified sphenethmoid, prootics, epipterygoids), the specimen represents one of the best examples of a markedly immature trematopid, an important data point for understanding the early ontogeny of trematopids. Specifically, it provides evidence that variation in otic notch structure can be ontogenetically influenced, not only among eucacopine dissorophids but also among trematopids. We provisionally refer the specimen to cf. Acheloma based on the presence of a denticulate vomerine ridge and other qualitative features. However, we emphasize that the taxonomic referral is complicated by several factors that more broadly confound trematopid taxonomy. This includes a low sample size (n = 1) of many taxa and marked size, and presumed ontogenetic, disparity between the known size range of different taxa. Complementary reexamination of both Acheloma cumminsi and Acheloma dunni as part of this study also reveals that the former possesses lateral exposures of palatal bones, the presence/absence of which was the only formal character that previously differentiated the two species, although other qualitative features (e.g., size of the internarial fontanelle) may differentiate these two species. With respect to OMNH 79318, the taxonomic referral is tentative because the specimen also shares many qualitative attributes with Phonerpeton pricei, a trematopid represented only by small-bodied, probably immature individuals. However, many of these shared features are likely to be influenced by ontogeny or size. The subsequent challenges that we encountered in our taxonomic referral suggest that ontogeny may be confounding taxonomy in both diagnoses and phylogenetic analyses of trematopids and emphasize the need for careful study of how this affects our understanding of trematopid intrarelationships. Summary for non-scientists This study describes the complete skull of a relatively small trematopid, a group of terrestrial temnospondyls from the Permo-Carboniferous that is most readily identified by having an unusually long nostril and snout. Even though this skull is complete and immaculately preserved, it was actually quite complicated to figure out which genus and species this specimen belonged to. There's only one trematopid known from the same site in Oklahoma, Acheloma dunni, but there's a lot of qualitative differences in aspects such as the profile of the skull and relative proportions. Conversely, our specimen is more similar in many respects to a geologically older trematopid from Texas, Phonerpeton pricei. Might this indicate the presence of two trematopids at the Richards Spur locality? Not so fast. Acheloma dunni is best known from much larger individuals than our specimen, whereas all of the specimens of P. pricei are about the same size or smaller to ours. So what if the apparent similarities to Phonerpeton and the differences from Acheloma are just size-related - in other words, at least these long-snouted trematopids go through development in a similar fashion and Phonerpeton captures only the early stages and Acheloma mostly captures the later stages? After doing a pretty thorough examination of all of the differences, there are strong correlations between size and disparities in anatomy. One feature in particular stood out as a difference between our specimen and Phonerpeton, namely these lateral exposures of the palate. In Acheloma, including both large and small individuals, these exposures poke out toward the side and don't form part of the margin of the orbit. In Phonerpeton and other trematopids, these exposures poke upwards and do form part of the margin of the orbit. Our specimen doesn't have either condition, at least not externally, but when you look internally through the CT data, at least one of the elements with a lateral exposure is expanding sideways to intercede between two other bones, not upwards towards the orbit. The direction of expansion appears to be quite reliable and suggests that this is a juvenile Acheloma, not an individual of Phonerpeton. Why's it all matter? Development is very poorly constrained in the fossil record. As a result, comparisons, either qualitatively or in a quantitative phylogenetic analysis can be biased by disparity in relative maturity of taxa; a juvenile of one taxon may look very different from an adult of another (apples vs. oranges), and these differences may not persist when comparing adults. This can in turn fundamentally affect our interpretations of evolutionary history and relationships, so a better understanding of development through documentation of small, presumably immature individuals is important. FAQWhat's a trematopid? Trematopids, named for the genus Trematops from Texas (which is now a junior synonym of Acheloma), are a group of modestly large terrestrial temnospondyls from the Permo-Carboniferous. They are most readily recognized for having a particular long nostril (see 'A' in the below figure, which in most trematopids is partially subdivided to produce a "keyhole-shaped" profile (alternatively you could consider it a filled-in 8 or the profile of a peanut in shell) and stupidly big heads for their body. What the large nose is for is not resolved by any means (e.g., Dilkes, 1993); a bone (the septomaxilla, abbreviated 'smx' in 'B' in the below figure) that frames the back end of the nasal capsule (the organ in the nose) is found at the middle of the nostril in trematopids. This indicates that the nasal capsule itself did not expand greatly relative to other temnospondyls, so the extra space must have served some other purpose. A salt gland (gets rid of excess salt) has been a recurring idea in the classical literature, but unsurprisingly, this is mostly found in animals living in or near salty bodies of water (e.g., sharks, marine iguanas), not amphibians inferred to be living in a diversity of terrestrial environments (and not on tropical islands either).
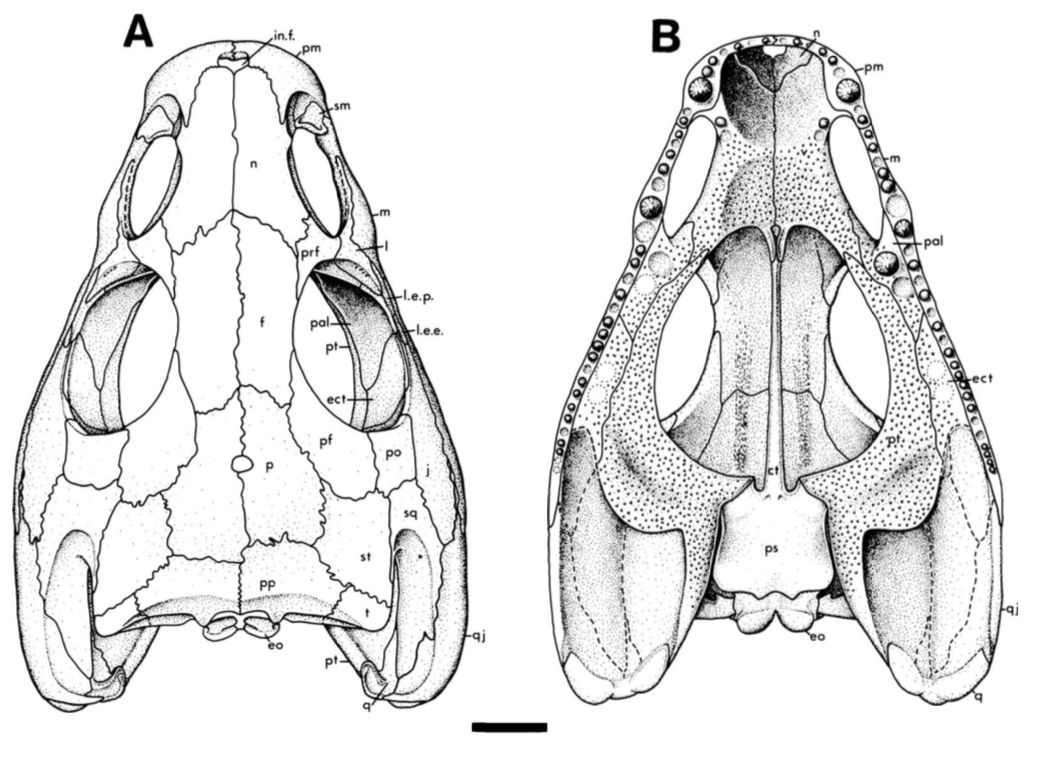
How so? The only trematopid that we have from this site (the Richards Spur locality that my lab works on) is Acheloma dunni (Polley & Reisz, 2011). The holotype of this species is huge (16.4 cm in length) compared to this little one that we described. Differences between the two specimens include some features that previous workers have suggested are useful for differentiating between trematopid taxa. For example, the otic notch of Acheloma is very narrowly constricted, unlike in this specimen and many other terrestrial temnospondyls in which is forms a 45-degree angle that is more open. The snout and the nostril are also proportionately shorter, and the skull profile doesn't quite look like that in Acheloma in which the skull really widens at the back end (see the below comparisons). Upon initial inspection, our trematopid doesn't really look like Acheloma. In fact, it looks a lot more like this one called Phonerpeton, from Texas.
So it's Phonerpeton? Not so fast. There's a catch. Phonerpeton is only known from small specimens, smaller in fact than our trematopid. What if the apparent greater degree of difference between our specimen and Acheloma is because of size? That would also account for why it looks a lot like the similarly-sized Phonerpeton. If you take a close look at our specimen, it's not very well-ossified. That is to say that its braincase is basically empty. No sphenethmoid, no prootic, small opisthotics, pretty open occiput...it's a long list. These are all features that we know characterize development in Acheloma dunni based on a study by Maddin et al. (2010) in which these features appear and become more developed in large individuals. The same lack of development is found in Phonerpeton! So it actually looks like Phonerpeton is a taxon represented only by juvenile individuals. At the very least, it hasn't been proven to be a diminutive trematopid. In short, a lot of this points to these differences between these really differently sized specimens being attributable to developmental (im)maturity, not the presence of two different taxa.
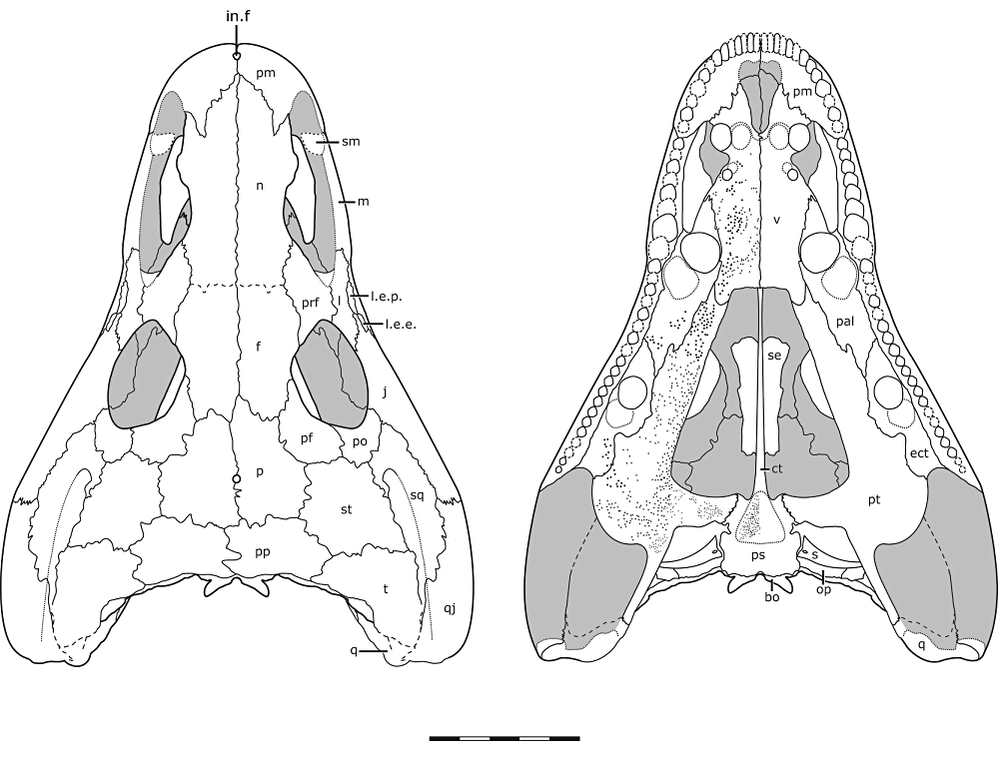
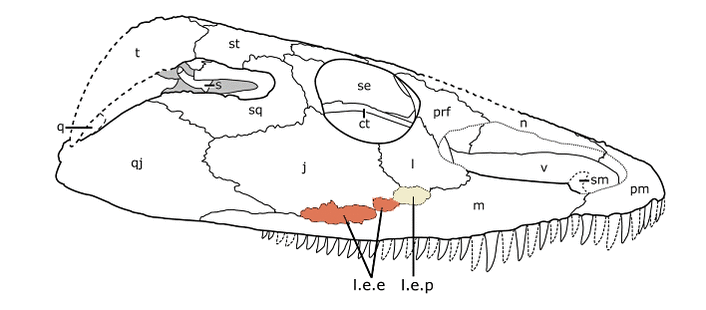
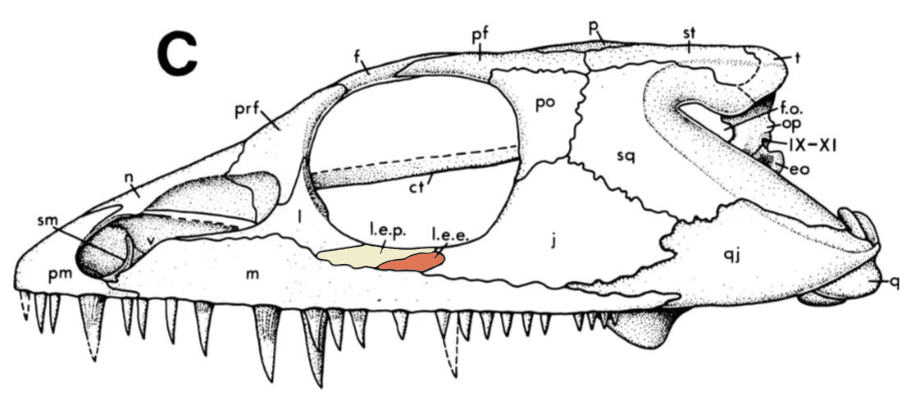
So...who is it.....? A key difference between Acheloma and all other trematopids is these things called lateral exposures of the palate (roof of the mouth) that are exposed on the outside of the skull. There are two, a lateral exposure of the palatine (LEP) and a lateral exposure of the ectopterygoid (LEE). In Acheloma, these exposures pop out between several other elements - basically, they don't enter the orbit (see Acheloma dunni below on left). In all other trematopids with the exposures, regardless of size, they enter the orbit (see Phonerpeton below on right). Seems pretty reliable to use to make a taxonomic distinction...
SO WHAT IS THIS ANIMAL?? After a lot of going around in circles and then up and down through some of the philosophical rabbit holes that I enjoy diving into, we ended up calling it cf. Acheloma, which basically means we think it's Acheloma, but we're not 100% sure. Read the paper if you want all of the gory details. What we are sure of is that this specimen is really immature, and that made for a lot of interesting talking points... And those are? Ontogeny can really mess with phylogenetic analyses to infer evolutionary relationships because characters that only track simple binary states like presence or absence of a certain element can't distinguish between absence that's phylogenetically informative (i.e. adult of taxon A has it, adult of taxon B doesn't have it) and absence that's ontogenetically informative (e.g., juvenile of taxon A doesn't have it, adult of taxon A develops it). Because most parts of the skeleton appear and become more developed throughout development, a really immature animal may appear to be more primitive from a phylogenetic standpoint (many features absent or underdeveloped). This can lead individuals of the same species but of different ages to be recovered as seemingly different species in a phylogenetic analysis because the analysis cannot differentiate between a condition that is the result of immaturity versus one that is the result of speciation. Taxa represented only by small individuals may appear more primitive than those only known from large individuals. This is obviously greatly misleading for our interpretations of evolution. Ontogeny can also mess with basic taxonomy, like figuring out which taxon a specimen belongs to (as in this situation). If a taxon is formally diagnosed based on features that only show up in large, presumably mature animals, then it would be difficult to confidently identify a juvenile that probably lacks many of them. The flip side is if a taxon is only diagnosed based on a small and probably immature individual, identifying adults may be equally challenging. These are a major issue for trematopids because some taxa are only known from small specimens and others are only known from large ones (Milner, 2018 has a good discussion of this). The smallest complete skulls are less than 5 cm (Rotaryus from the early Permian of Germany), while the largest are 18 cm (Acheloma from the early Permian of Texas). Even if they didn't all reach the same adult size, this is a giant difference in size and presumed developmental maturity, making any phylogenetic analysis susceptible to the above problems. A number of features also correlate pretty well with size in trematopids, such as an open or closed otic notch (closed only in large ones) and greatly elongated snouts (seen best in large specimens of Acheloma). Basically, this makes it really challenging to be confident in an analysis that broadly samples a lot of presumably immature taxa. Anything else cool? You can only see this if you go to the supplemental, but it looks like this trematopid might not have finished eating dinner when it died! I'm not joking either. There's the pelvis and upper leg of a smaller temnospondyl sitting in the mouth of our trematopid (see the black rectangle in 'A'). Whether it was actually dinner or just ended up in the mouth through serendipity is impossible to prove, but one would certainly predict that this trematopid was munching on its distant cousins! 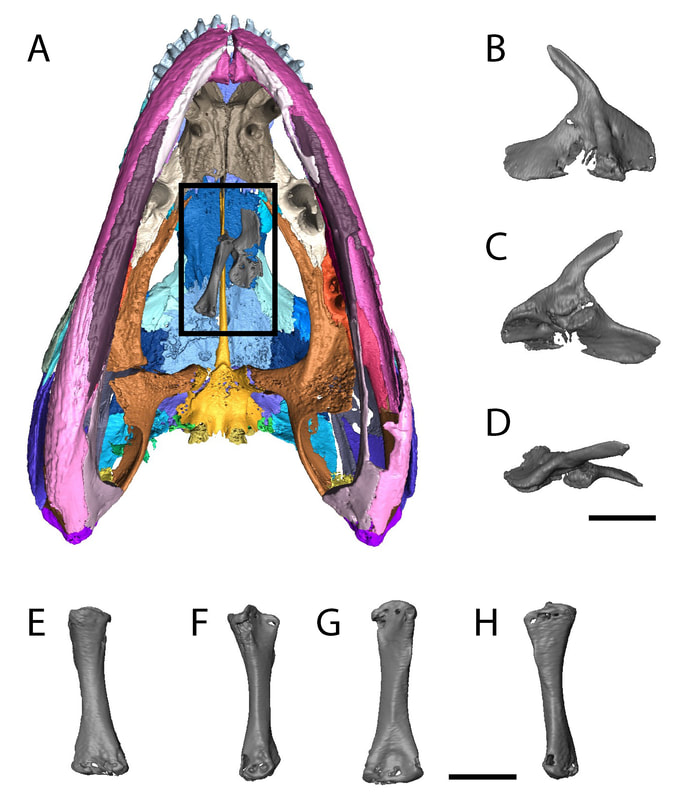 Refs
Comments are closed.
|
About the blogA blog on all things temnospondyl written by someone who spends too much time thinking about them. Covers all aspects of temnospondyl paleobiology and ongoing research (not just mine). Categories
All
Archives
January 2024
|
 RSS Feed
RSS Feed
