One of the above is a latrine. You may have observed one over the course of your life. The other one is an extinct amphibian. You probably have not observed one over the course of your life. "Toiletheads" and "toilet seat mechanism" are widely used colloquialisms to refer to temnospondyls because of a perception that their mouths opened like a toilet seat, namely by the skull (= the lid) raising up, with little to no movement of the jaw (= the seat). This is in contrast to most other jawed vertebrates, which like us, primarily depress (lower) the jaw to open our mouths. This peculiar attribute of temnospondyls is probably one of the most widely disseminated facts about them among the broader paleontological community. As evidenced by my popular Valentine's rhyme from last week, people think this is hilarious. It makes temnospondyls relatable. However, most people (scientists or otherwise) don't actually know where this idea comes from, how well it's been tested, or how broadly it may apply within Temnospondyli. So, I thought that I would spend this week going over the evidence, the lack of evidence, and how to interpret it.
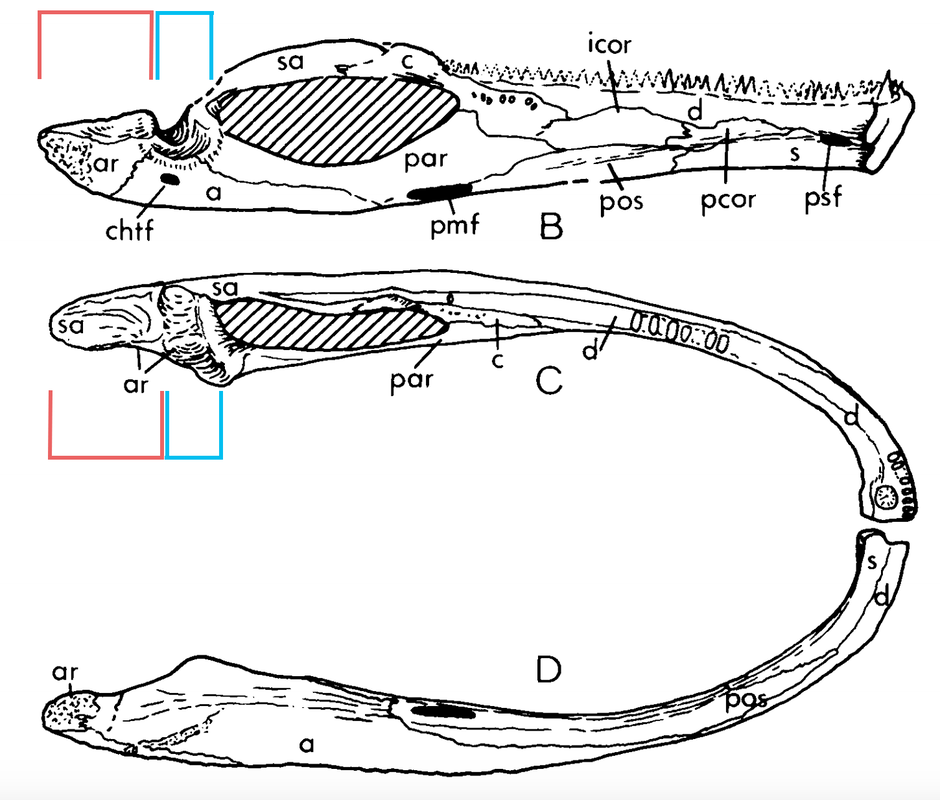
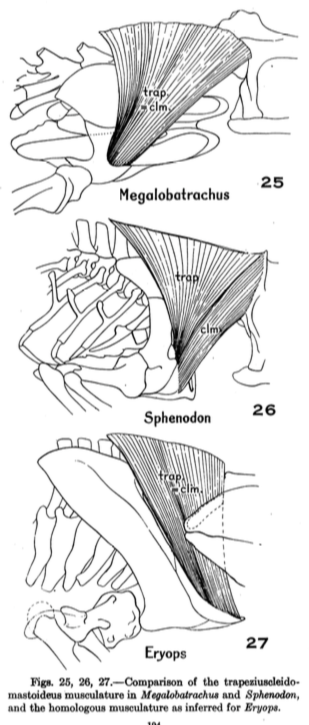
Watson reasoned that the relative position of the skull-neck joint to the skull-jaw joint was an important consideration. If the skull-neck joint was well in front of the skull-jaw joint, the animal would have to lift up the front part of its body in order to raise the skull to open its mouth. Conversely, if the two joints were in about the same transverse plane (the same longitudinal position), then the skull could be raised without having to lift the body. Watson also argued that such a method of opening the mouth explained two features of "flattened" temnospondyls: (1) the development of the paired exoccipital condyles and (2) the gradual development of a long process at the back end of the jaw, called the retroarticular process. Because he inferred that the flat skull would have created a mechanical disadvantage for the neck muscles projecting from the back of the skull, he reasoned that the attachment of another muscle (the depressor mandibulae) to the long posterior process would have created a compensatory mechanical advantage. The large condyles would then have permitted great dorsoventral (up and down) flexibility at the skull-neck joint, and the jaw would have moved forward as the mouth opened. This idea was expanded on in later years by Watson (e.g., 1958, 1962) for a number of well-known temnospondyls, always with a focus on the depressor mandibulae as the primary jaw-opening muscle. A few subsequent workers of the mid-20th century (e.g., Panchen, 1959) extended this model to other groups, retaining the depressor mandibulae as the primary (but not only) muscle involved with this motion.
Howie (1970) was the first worker to propose a major departure from Watson's original model. Howie utilized brachyopids, whose skull-neck joint is well behind the skull-jaw joint, in contrast to other flat-skulled temnospondyls. In order for the depressor mandibulae to function in the way that Watson proposed, the skull-neck joint (and with it, the vertebral column) would have to be displaced downward by a substantial amount, implying that either (a) the vertebral column was very loosely attached to the shoulder girdle, allowing it to be greatly displaced or (b) the shoulder girdle was not in contact with the ground, an unlikely idea if the jaw had to remain in contact with the ground to act as a fulcrum (think of it as planking with your chin on the ground). This also raises the problem of how could the jaw move forward during a gape if if was needed as a fulcrum.
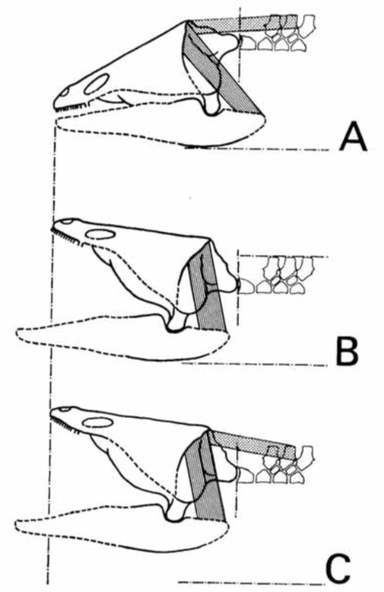
Howie also pointed out that Watson's idea of a depressor attached to the top of the back of the skull was infeasible because of how the hypothesized angle of the muscle would have resulted in opposing forces, a lateral (outward) rotation around the skull-jaw joint with a medial (inward) pull of the depressor. Howie thus proposed a different model (part B on the above left figure), in which two main muscle groups were involved.
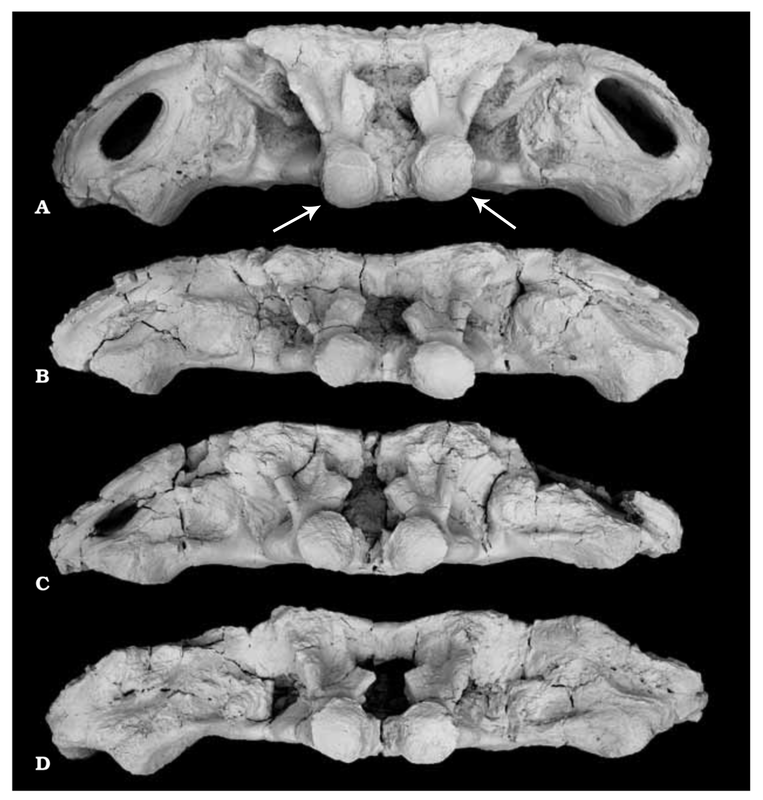
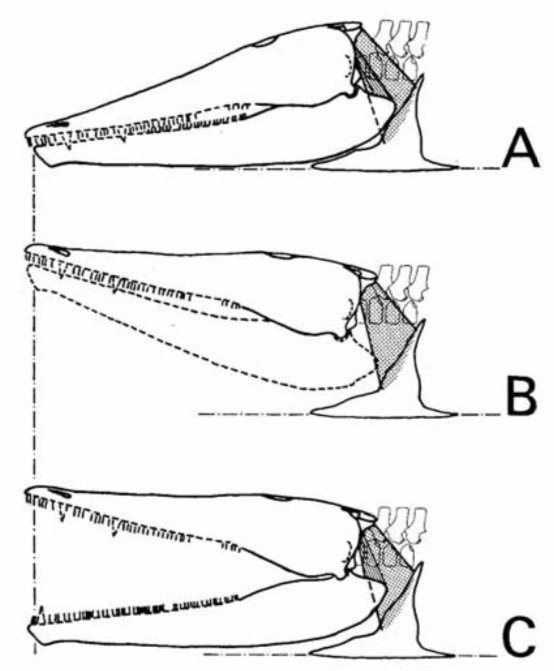
Howie also addressed the role of the retroarticular process. Temnospondyls with a more sloped occiput, like brachyopids, would need a longer retroarticular process. However, an alternative solution would be to have an upturned process that could produce the same mechanical advantage, but without needing to be extensively long. The overall morphology of the jaw articulation probably constrained the gape (degree of opening of the mouth) quite substantially. In particular, the relationship of the quadratojugals to the quadrate (the latter being the skull bone that forms the skull-jaw articulation) would limit the lateral movement of the jaws during opening, which was a result of the morphology of the quadrate condyles (the part that contacts the jaw). Another limitation may have been the position of the orbits; temnospondyls with the eyes far back on the skull would probably have major issues actually seeing much of anything if the skull was raised too far, even if the eyes protruded above the skull to look forward. Subsequent workers have generally proposed gapes around 20-30° (e.g., Shishkin, 1987; Hellrung, 2003), although plenty of other angles (up to a right angle) have been suggested as well. 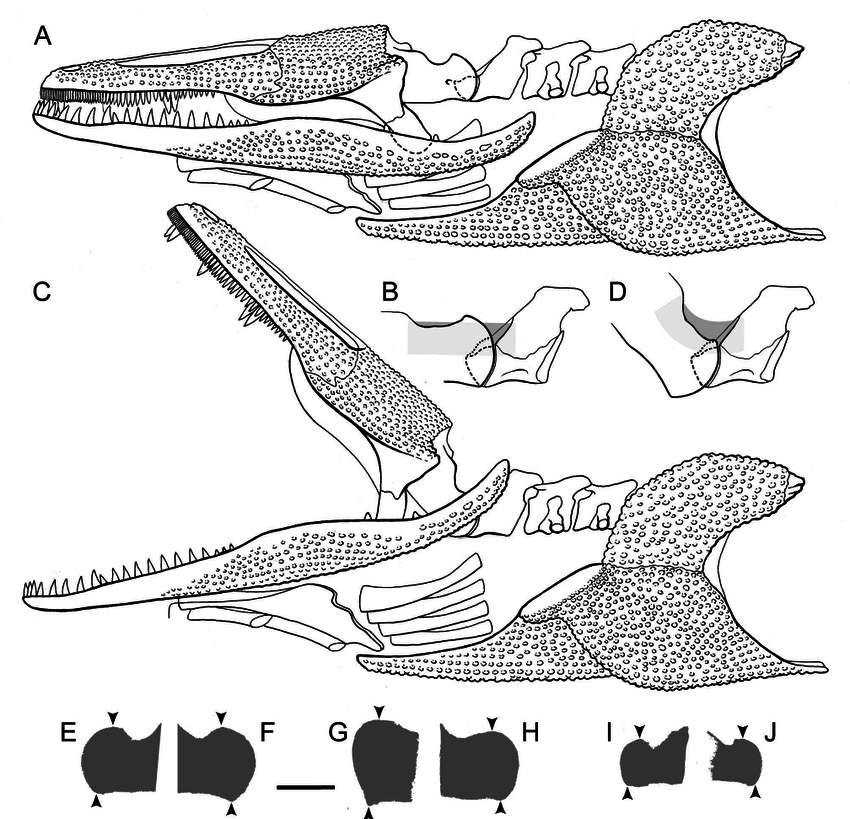 Model of head raising in Gerrothorax pulcherrimus. (A-B) resting state; (C-D) gaping state. A and C show the entire skull, mandible, and pectoral girdle, whereas B and D show the skull-neck (atlanto-occipital) joint. (E-J) silhouettes of the curvature of the condyles of three different specimens in medial (internal) view, showing the asymmetry of the dorsal and ventral contours; arrows indicating the margins of the articulating surfaces. Figure from Jenkins et al. (2008). Work by Jenkins et al. (2008) is the most recent and the most detailed work to explore models of head raising in temnospondyls, using the plagiosaurid Gerrothorax pulcherrimus.
A lot of the discussion on head raising is inherently linked to other aspects of feeding in stereospondyls. Unfortunately, a lot of that remains poorly resolved or tested only in very specific and isolated groups. For example, were most stereospondyls active swimmers and hunters in the water column (pelagic) or did they prefer ambush tactics at the bottom (benthic)? Did they directly bite prey or attempt to inhale them through suction feeding? Watson's classical model of head raising was predicated on the assumption that the animal sat on the bottom so that its jaw was firmly in contact with the ground and could thus be used as a fulcrum, although a general bottom-dwelling ecology is not incompatible with the more modern concepts of head raising. There has been other work done on feeding in temnospondyls, primarily finite element analyses (FEA) by Fortuny and colleagues and reconstructions of muscles systems by Witzmann and colleagues, but these usually are usually intended to assess other aspects of temnospondyl feeding and do not directly inform on the viability and degree of head raising. Some examples:
What's it all mean? Okay, so now that we've gone over all of that, let's sum it all up and debunk some myths and misconceptions:
Refs
2/22/2019 09:20:51 am
Hi Bryan,
David Marjanović
2/22/2019 10:34:01 am
*Koolasuchus* is Cretaceous, not Jurassic! Comments are closed.
|
About the blogA blog on all things temnospondyl written by someone who spends too much time thinking about them. Covers all aspects of temnospondyl paleobiology and ongoing research (not just mine). Categories
All
Archives
January 2024
|


 RSS Feed
RSS Feed
