|
Cacops and Dissorophus might be the best-represented dissorophids in the scientific literature, but neither is the most famous dissorophid. That honour falls to Platyhystrix (pronounced 'plat-e-his-tricks'), the "flat porcupine." Sure, this one's never made it into any movies, but neither have practically any temnospondyls. Darren Naish did a Tet Zoo blog post on Platyhystrix a few years back, and the internet is rife with reconstructions (of varying quality and accuracy) of this peculiar animal. There's even a plastic toy model (1:8 scale) that was made by Play Visions as part of a 1998 extinct amphibian series that included Diplocaulus, Peltobatrachus, and Gerrothorax (this is apparently a highly valued and rare collector's set that I may have had at some point in my childhood). It has yet to make it into one of the mobile Jurassic World games, arguably the "I know I've made it when..." but also the pinnacle of "....that is not what I look like." But how much of Platyhystrix do we actually know, and why it is so popular?
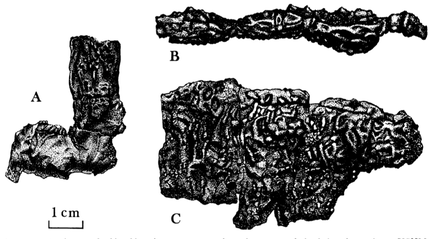
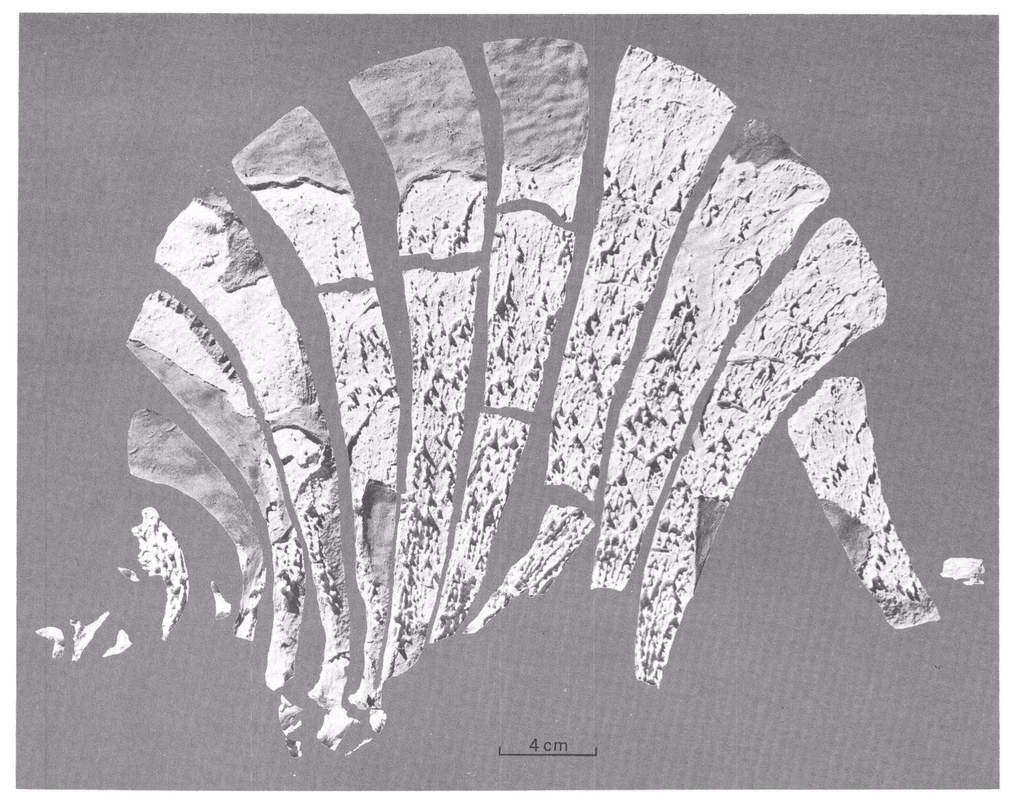
 Photograph of some of the holotype of Platyhystrix rugosus at the American Museum of Natural History. (C) Bryan Gee. Photograph of some of the holotype of Platyhystrix rugosus at the American Museum of Natural History. (C) Bryan Gee. A (not so) brief history of Platyhystrix The material now regarded as the holotype of Platyhystrix rugosus consists of only a few fragmentary neural spines from the early Permian Cutler Formation of New Mexico. It was first referred to "Zatrachys apicalis" by American palaeontologist E.D. Cope, a taxon that was described in 1881. It gets kind of confusing because the specimen, AMNH FARB 4785, is the number of the type specimen of "Z. apicalis." However, the holotype of P. rugosus was extracted from this specimen but not given a new number. The non-Platyhystrix component of the specimen is now regarded as some indeterminate aspidosaurine and was apparently lost at some point. The Platyhystrix component remains, with the same number, as a type specimen. This component was then placed within a new genus by E.C. Case (1910), who argued that it was the same as the Early Triassic poposauroid archosaur "Ctenosaurus" (now Ctenosauriscus), which was at the time thought to be a relict pelycosaurian synapsid. S.W. Williston (1911) recognized that the material was not "Ctenosaurus" and thus created the genus Platyhystrix, but he considered it rather unlikely that it was an amphibian, even though he noted that an amphibian was his initial inkling based on the similarity of the neural arch to that of Eryops. Williston (1916) then placed it among the temnospondyls based on some fragmentary skull material that appears doubtfully referred since it has prominent spike-like projections that are more like that seen in zatracheids (not dissorophoids but perhaps closely related). Unsurprisingly, the exact taxonomic affinities of Platyhystrix remained poorly agreed upon for several decades. Wann Langston (1953) described some additional postcranial fragments and a handful of cranial fragments with similar ornamentation and discussed the history to date of the taxon; he considered it to be a close relative of the eryopoids. Other workers (e.g., Carroll, 1964; DeMar, 1968) considered it to be either closely related to the dissorophids or to be within the dissorophids. In the meanwhile, fragmentary bits of Platyhystrix continued to turn up in the Four Corners states, essentially restricted to the unique neural spines since that was the only diagnostic skeletal feature (e.g., Vaughn, 1962, 1963; Carroll, 1964). This is of course one of the big problems with naming new taxa based on very fragmentary material: it can be near impossible to prove that a more complete specimen belongs to that taxon or to confidently exclude it from being that taxon. In this case, it was essentially impossible to identify any cranial material of Platyhystrix unless it came with a bunch of clearly associated neural spines - an association that is fairly rare in the fossil record. It was thus not until the work of Berman et al. (1981), who described one such specimen that was collected 100 years prior, that the cranial anatomy became appreciably well-known. This solidified its placement within dissorophids, where it has remained ever since. Subsequent workers have reported more fragmentary neural spines (e.g., Lucas et al., 2005; Huttenlocker et al., 2018). There is only one recognized species: Platyhystrix rugosus.
What makes Platyhystrix special? Platyhystrix is best-known for its greatly elongated neural spines, which are reminiscent of those found in early synapsids like Dimetrodon and Edaphosaurus. Below is one of the best-known specimens of these spines, a natural mould on the left and the cast that was made from it on the right (figures from Lewis & Vaughn, 1965). It's assumed that this was the natural arrangement of the spines - in close position but not contact with each other and forming a smoothly convex dorsal margin. In the figure on the right, the head would have been to the right. As you can see, the spines are essentially never perfectly vertical; they always curve slightly in one direction or another, and this is stronger toward the pelvis (to the left). On the flip side, there isn't as much that can be said about the skull because of how poorly known and poorly preserved it is. Some parts do appear to have a very developed nodular (papillose) ridge-like development in some areas that at least hasn't been reported from other dissorophids.
 Illustrations of the holotype of Astreptorhachis ohioensis. A and C are lateral views of two different fragments, and B is the dorsal view of the same specimen as in C. Figure from Vaughn (1971). Illustrations of the holotype of Astreptorhachis ohioensis. A and C are lateral views of two different fragments, and B is the dorsal view of the same specimen as in C. Figure from Vaughn (1971). What's up with that sail? The short answer is that there is no conclusive answer. Sails have been far more extensively explored in synapsids like Dimetrodon (e.g., Bennett, 1996; Huttenlocker et al., 2010, 2011; Rega et al., 2012) and to a lesser degree in spinosaurid dinosaurs (e.g., Gimsa et al., 2016; Henderson, 2018). DeMar (1966) made a detailed synthesis of the morphology and diversity of dissorophid osteoderms, but Platyhystrix was not regarded as an unequivocal dissorophid at the time, and it was thus only mentioned very briefly in passing as being somewhat similar to Aspidosaurus. It's very interesting to note that (1) hyperelongation of spines occurs in several dissorophids and (2) occurs largely in Carboniferous taxa. Vaughn (1971) reported a new dissorophid from the Carboniferous of Ohio, Astreptorhachis ohioensis, that has not only hyperelongation of the spines but also apparent fusion between successive neural arches. There is also a presently undescribed dissorophid from the Carboniferous of Nova Scotia that exhibits similar features. One species of Aspidosaurus, the Permian-age Aspidosaurus binasser, is reported to have hyperelongate spines in part of the axial column. Vaughn (1971) speculated that the spines primarily served to stiffen the trunk as part of the suite of terrestrial adaptations of dissorophids; this would be comparable to the proposed primary function of the osteoderms in other taxa (Dilkes & Brown, 2007; Dilkes, 2009). Does Platyhystrix have osteoderms? Generally speaking, it is considered to only have ornamentation on the spines, not distinct osteoderms (e.g., Vaughn, 1971; Schoch & Milner, 2014; but see Witzmann & Soler‐Gijón, 2010 for another perspective). Whether the ornamentation of neural spines is the intermediate step that produced the osteoderms of more derived dissorophids remains unclear. The function of hyperelongation in dissorophids hardly need be related in any fashion to that of amniotes, but the same hypotheses (e.g., thermoregulation, sexual dimorphism, species identification, etc.) should be considered until they can be more definitively tested. Refs
Comments are closed.
|
About the blogA blog on all things temnospondyl written by someone who spends too much time thinking about them. Covers all aspects of temnospondyl paleobiology and ongoing research (not just mine). Categories
All
Archives
January 2024
|


 RSS Feed
RSS Feed
