|
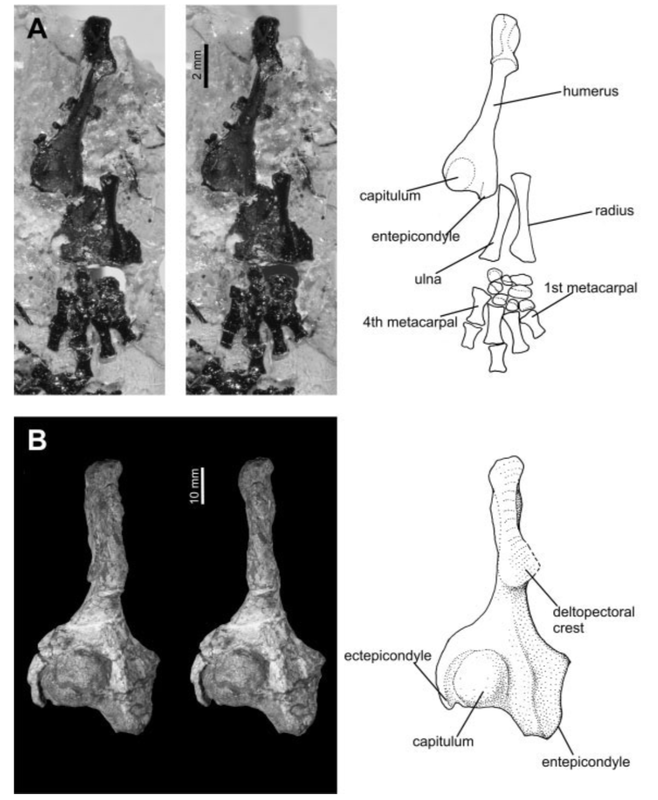
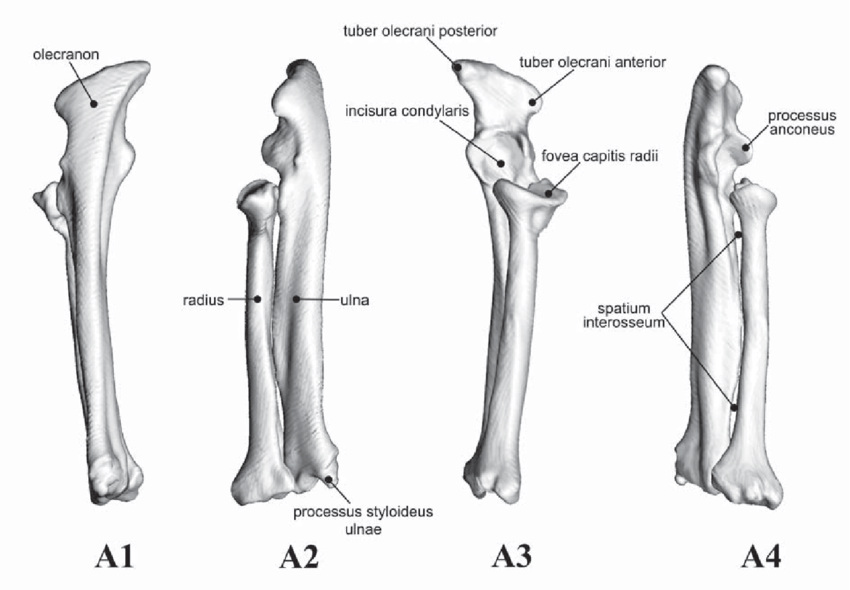
Last week, I covered what the skull might tell us about an animal's ecology. The skull contains a lot of important sensory organs and is a major part of several major functions, like feeding and breathing, not to mention that skulls are very diagnostic in the fossil record compared to other elements. However, one important function that the skull typically does not fulfill is locomotion (the exception is animals that use their head to burrow into dirt or sand). Because locomotion is such a critical function, tetrapods have modified their limbs in a remarkable variety of ways to adapt to their local environment. Fundamentally, the same bones are present in the limbs of all tetrapods, but different ones have taken on drastically different shapes to best serve their function. For example, the limbs of swimming mammals (particularly the arms) have shortened drastically, while the fingers have become elongated, forming the distinctive paddle. 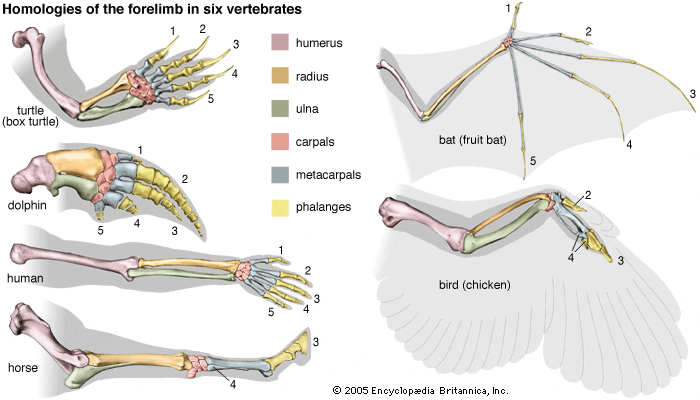 Arguably, the limb might be the most informative singular feature that can inform the lifestyle of extinct tetrapods since we can't observe their locomotion itself. There are very clear suites of changes that occur when animals adapt to different lifestyles, such as climbing (scansorial), burrowing (fossorial), or swimming (natatorial), many of which can be seen in the above diagram which depicts a variety of different tetrapods that encompass different lifestyles on land, in the water, or both. 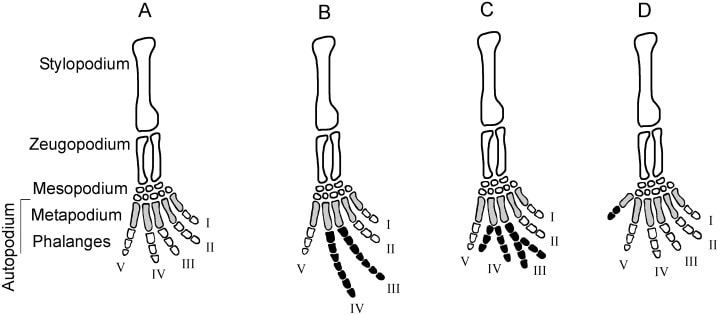 The limb is divided into several regions with fancy names that correspond to less fancy names that normal people use. The stylopodium is the upper limb bone on either the arm (humerus) or the leg (femur). This is what articulates with your shoulder, and it will always be a single bone. The zeugopodium is the lower limb bones on either the arm (radius, ulna) or the leg (tibia, fibula). The intersection between the stylopodium and the zeugopodium forms the elbow joint. Usually these bones are paired, but in some instances, they can end up fusing together, which is what happens in living frogs as an adaptation for their jumping mode of locomotion (saltation). In frogs, these composite bones are named exactly like you would expect: the radius + the ulna = the radioulna, and the tibia + the fibula = the tibiofibula.
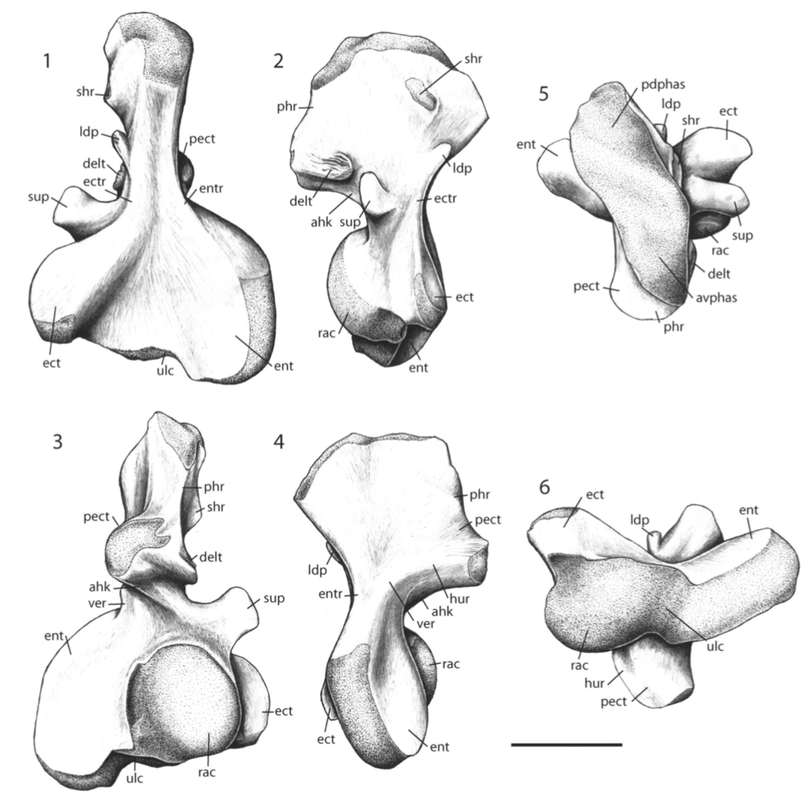
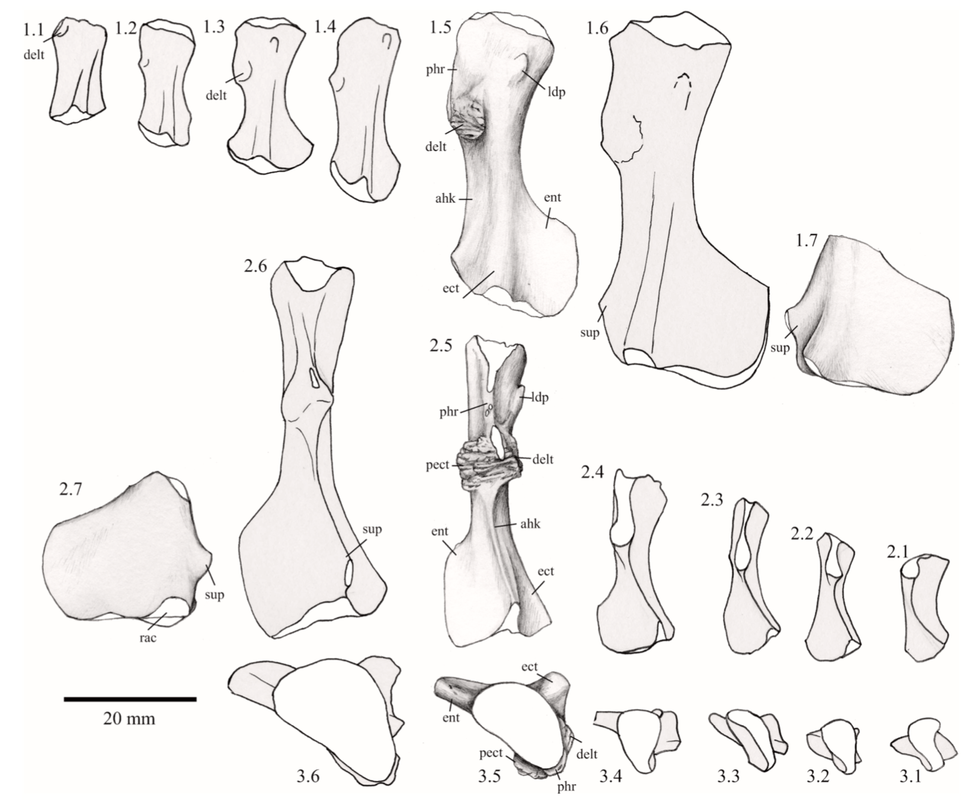
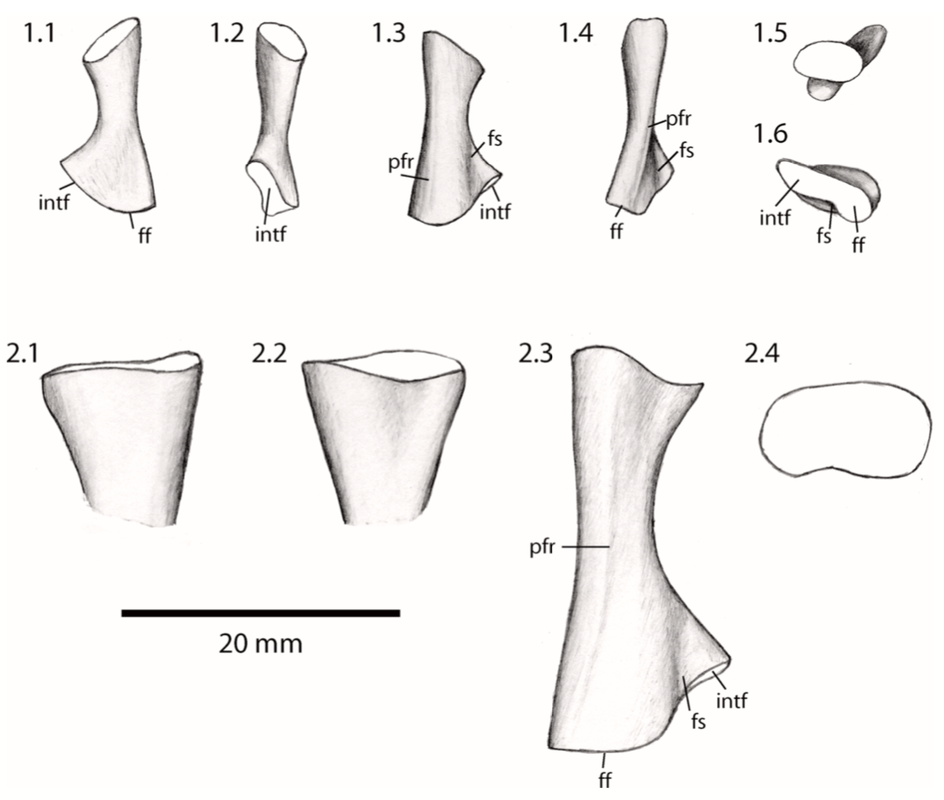
Humour the humerus The humerus, or the upper arm bone, is often the butt of various puns (not just among scientists), but its etymology is strictly anatomical in nature despite the phonetic convergence. The humerus is one of the most morphologically diverse elements of the limb, in part because some animals put weight on it (quadrapeds, walk on four legs) but others like humans do no (bipeds, walk on two legs). This has freed up the humerus to be adapted to suit quite a lot of functions associated with the forelimb, some of which mostly happen downstream (e.g., grasping branches) but because the humerus is the junction between the shoulder and the hand, it serves as a pivotal feature in the tetrapod skeleton.
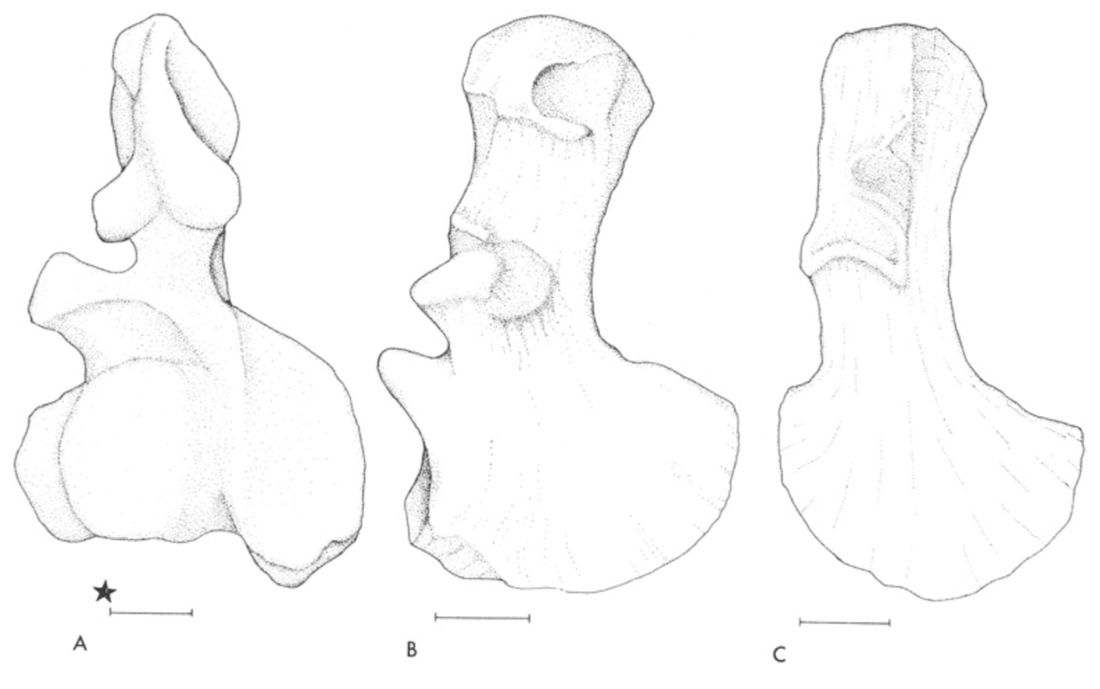
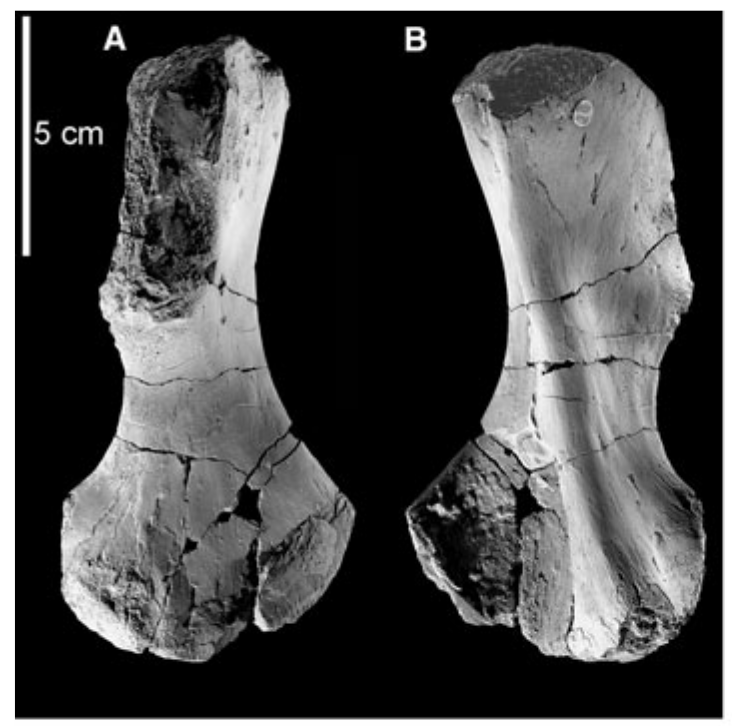
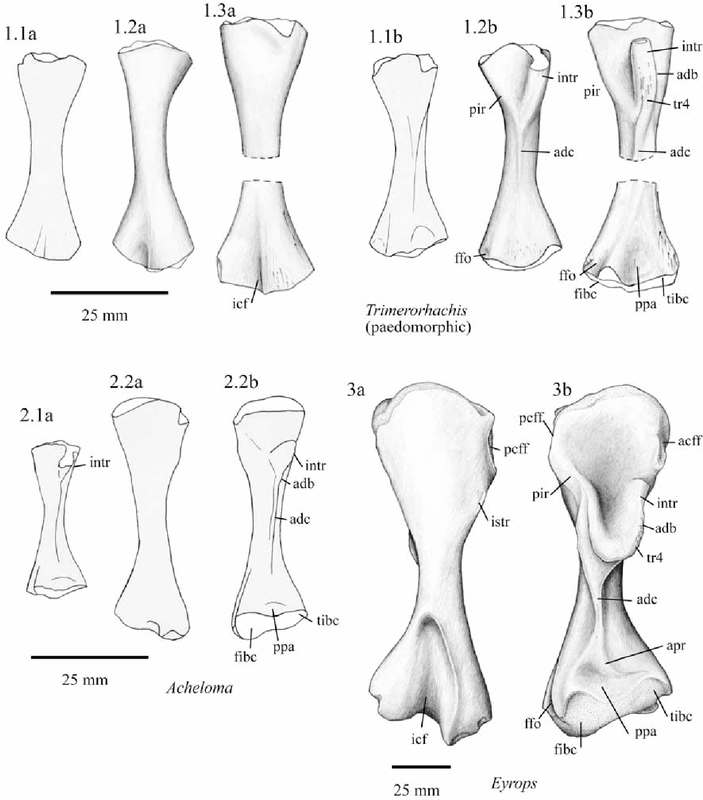
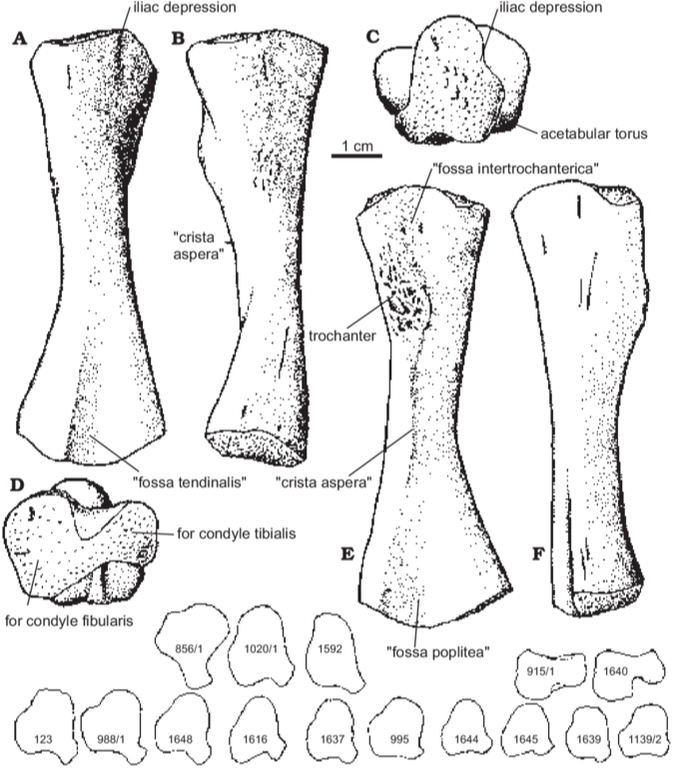
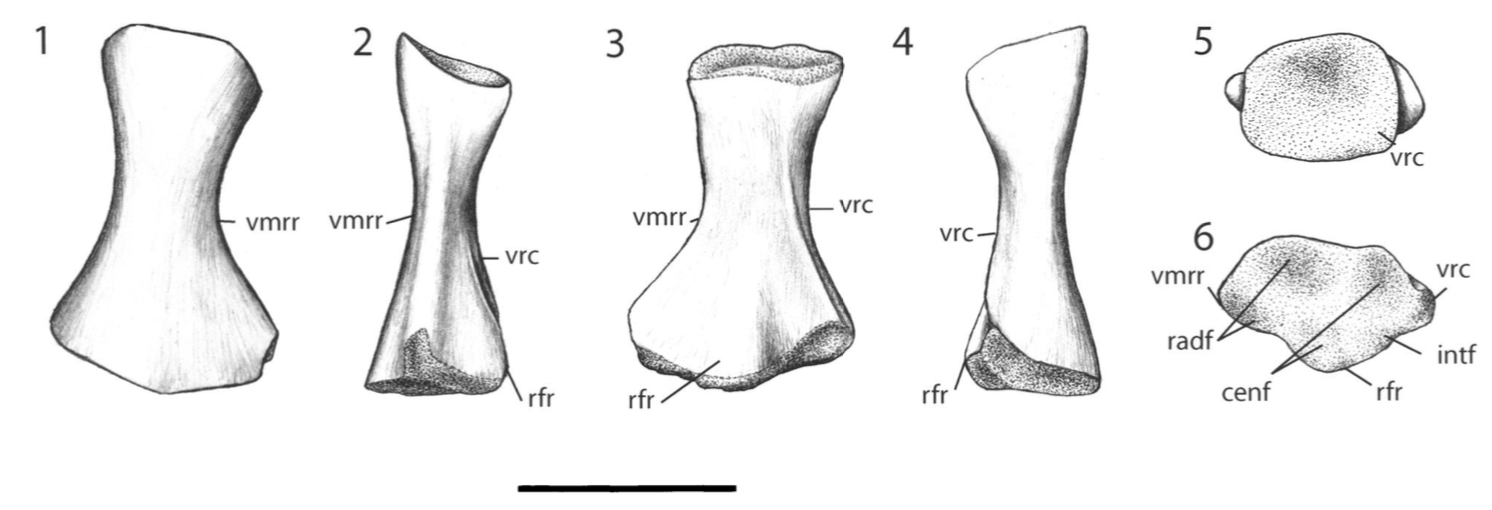
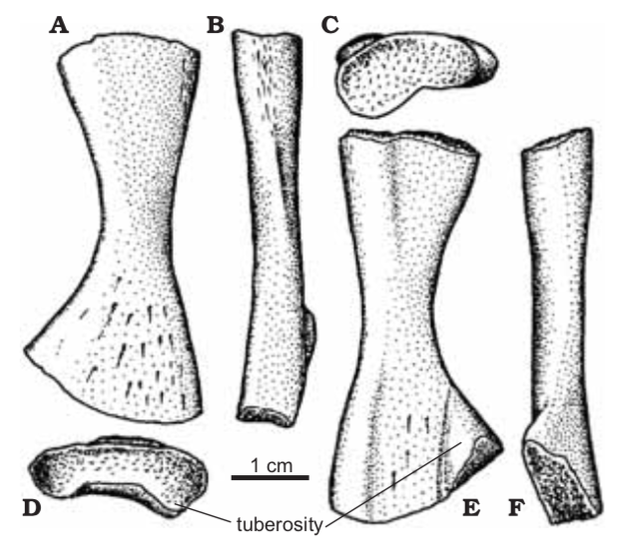
Proportions and degree of development are the two most distinctive ways to differentiate the humerus. Longer and more slender humeri, like that of Doleserpeton above on the right, are taken as evidence for a terrestrial lifestyle. Terrestrial animals tend to have longer limbs, as there are many advantages (e.g., longer running stride, increased height), whereas these advantages don't translate to the aquatic realm, in which the drag of moving in water often leads the humerus and other bones to actually become proportionately shorter. The humeri of the metoposaurid Anaschisma (above on the top left, part C) and the capitosaur Cyclotosaurus (above on the bottom left) are a classic example of a short and stocky humerus that most workers agree is evidence for an aquatic lifestyle. The angle at which the two ends are set is also used as a predicting factor. If the heads are set at nearly a right angle to each other (highly torqued), like in Doleserpeton and Dissorophus, this is taken as evidence for a terrestrial lifestyle. Something closer to 70 degrees is taken as evidence for an aquatic to semi-aquatic lifestyle. Without going into the nitty gritty of it, a higher degree of torsion stabilizes the elbow and increases the stride.
Finally, the development of different attachment sites for muscles, such as the supinator process (which may not actually be a real supinator process; Bishop, 2014) and the deltopectoral crest (these are abbreviated 'sup' and 'delt' in the figures above), tend to be more developed in terrestrial temnospondyls due to the biomechanical demands on holding the body upright and then moving about on land (like Eryops above on left). A more aquatically inclined animal will have greatly underdeveloped attachment sites and processes or none at all (like Trimerorhachis above on right). Note, however, that any single site/process (or feature in general) is not always a good proxy on its own; some temnospondyls, like dissorophids, which otherwise exhibit all of the hallmarks of being very capable on land, lack a supinator process. It is sometimes easier to just consider things in aggregate in terms of how "complex" they look. A second example is the torsion observed in Trimerorhachis; there is considerable variation from about 45 degrees to 90 degrees but no clear pattern (e.g., increasing torsion with increasing size) or explanation. Femur
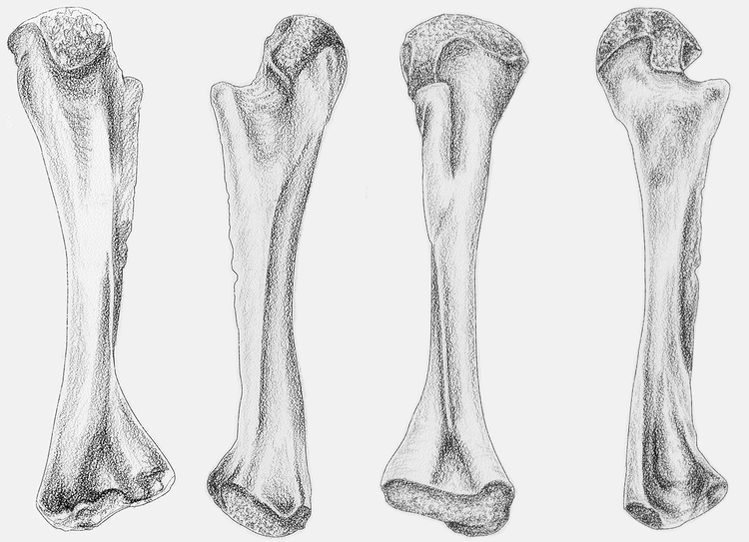
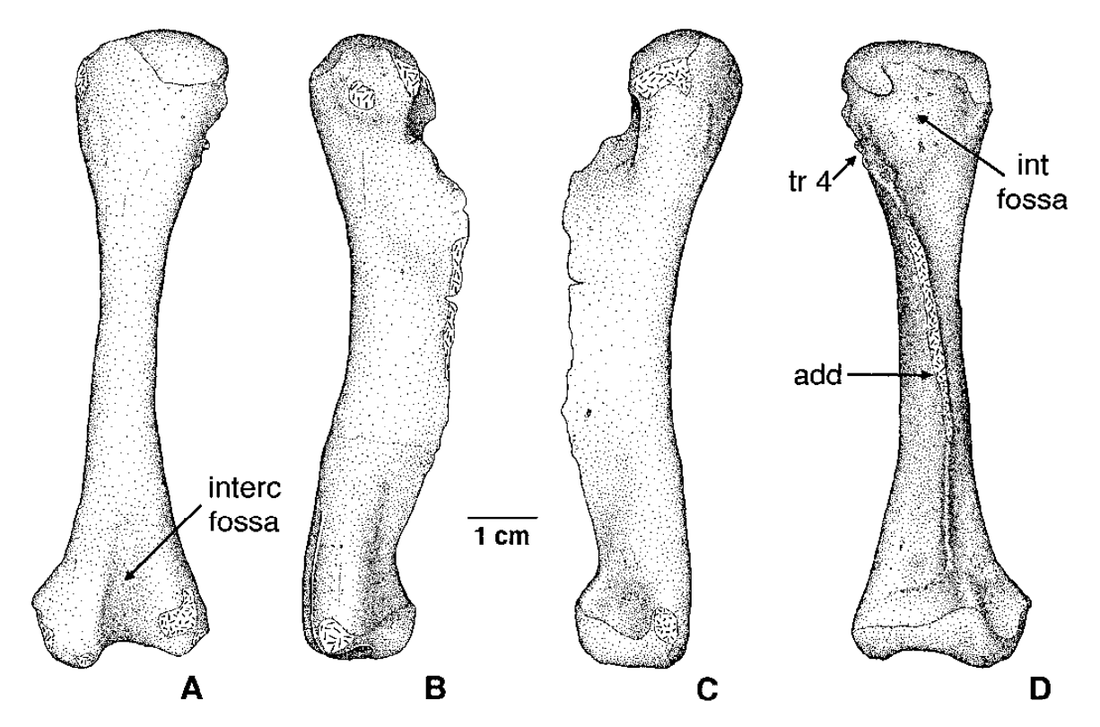
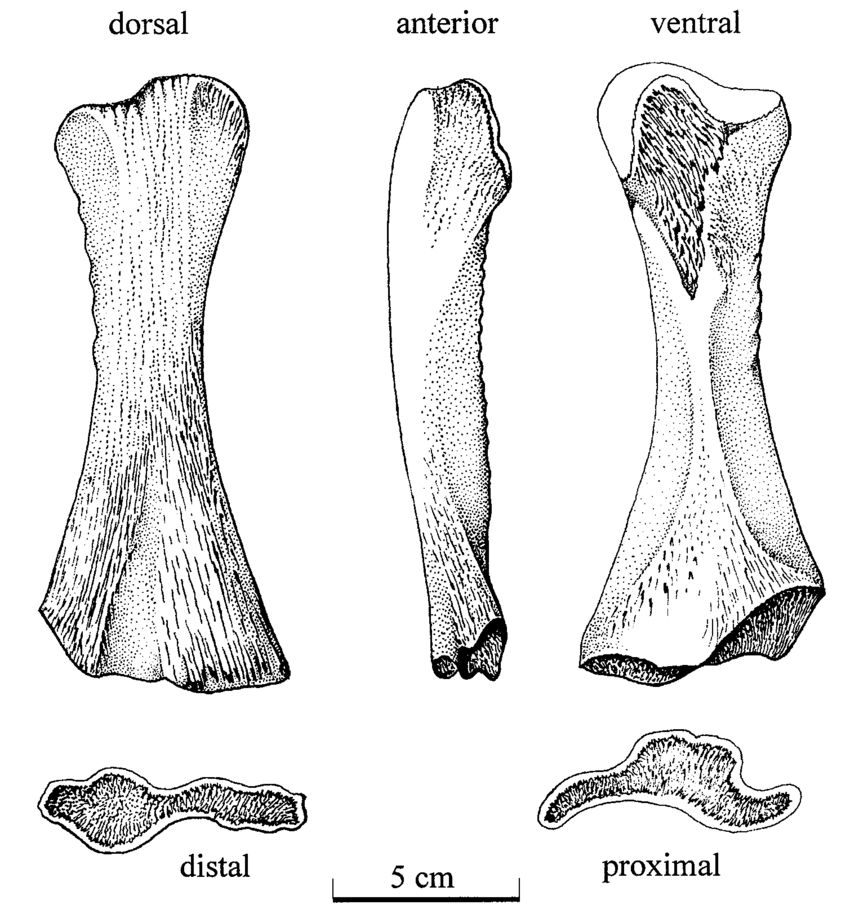
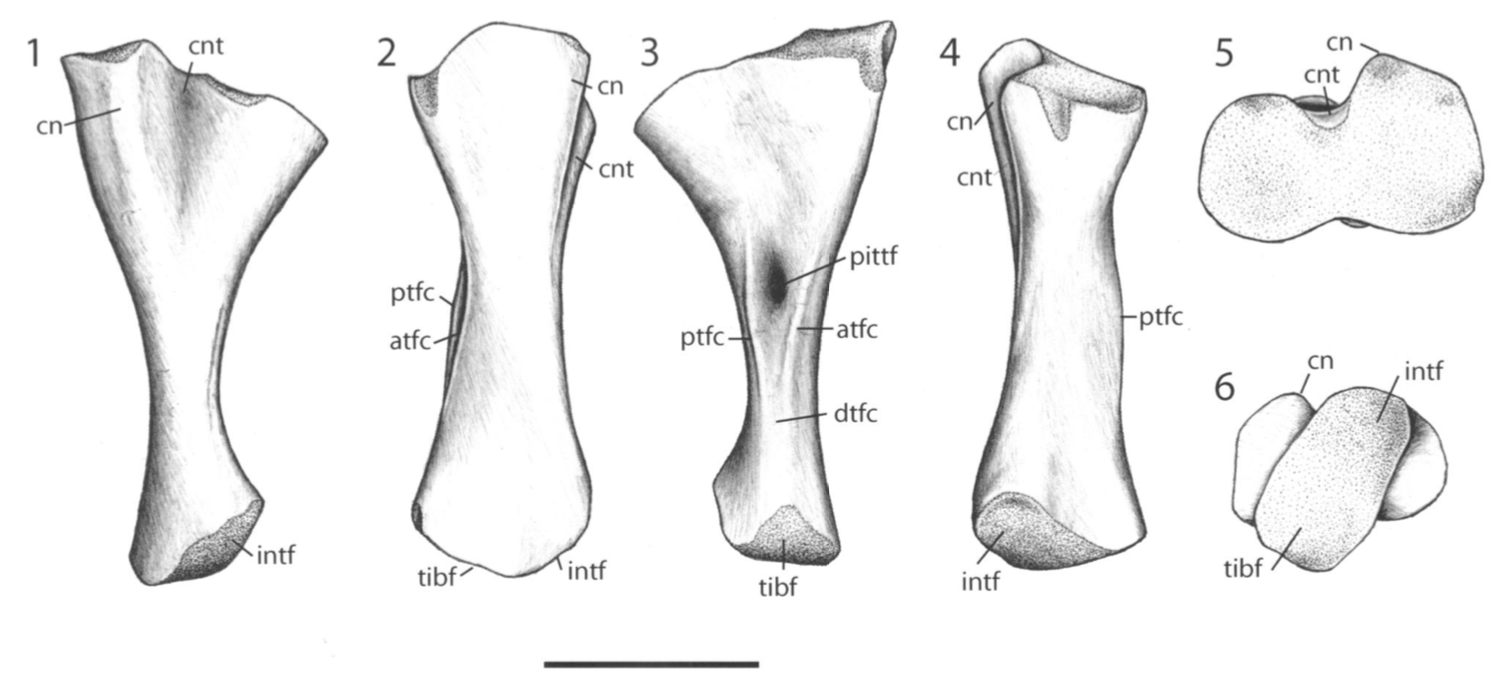
Terrestrially inclined The degree to which the ends are ossified is true of all limb bones, but it tends to be the most obvious in the femur because a poorly ossified femur will be squared-off / flat on the ends, and a well-ossified femur will be convex on both ends, with a distinct pair of rounded ball-like condyles for the tibia and fibula at the "bottom" end (the distal end, or where it meets the ankle). Really well-ossified surfaces are found in terrestrial temnospondyls like Doleserpeton (above on left) and Cacops (above on right). Typically, we then assume that poorly ossified femora belong to aquatic animals, like Mastodonsaurus (below on left) and Metoposaurus (below on right). However, poorly ossified femora can indicate not only a more aquatically inclined animal, but also an immature one (across the aquatic-terrestrial spectrum). This puts a real premium on having a good idea (and solid rationale) for inferring the relative age of a given specimen, which can end up being a very circular exercise when only using external anatomy.
Aquatically inclined Like the humerus, the degree of "development" of muscle attachment sites can also be insightful, although this too can be confounded by immaturity or other nuances of development. Examples for the femur include what's usually called the adductor crest (for the adductor muscles), a long blade-like ridge running down the length of the femur (the more it sticks out, the more terrestrial the taxon probably is); the internal trochanter (a variably developed thickening at one end of the adductor crest; a distinct process rather than a rugose patch of texture tends to indicate terrestriality); and the intertrochanteric fossa (a depression next to the internal trochanter; the deeper it is, the more musculature it's holding). Revelations of the radius The radius is the most generic looking limb bone out there. In pretty much all temnospondyls, it is just a cylindrical rod. These are sometimes confused for ribs, phalanges (finger bones), parts of the gill apparatus, and even the stapes (an ear bone). Sometimes there is a ridge on one surface that can help you ID it, but even that appears to only show up distinctly in adults. They are just that undiagnostic. Other than proportional differences that would be present in other limb bones, the radius has never proven to be particularly distinctive or informative for inferring lifestyle or ecology.
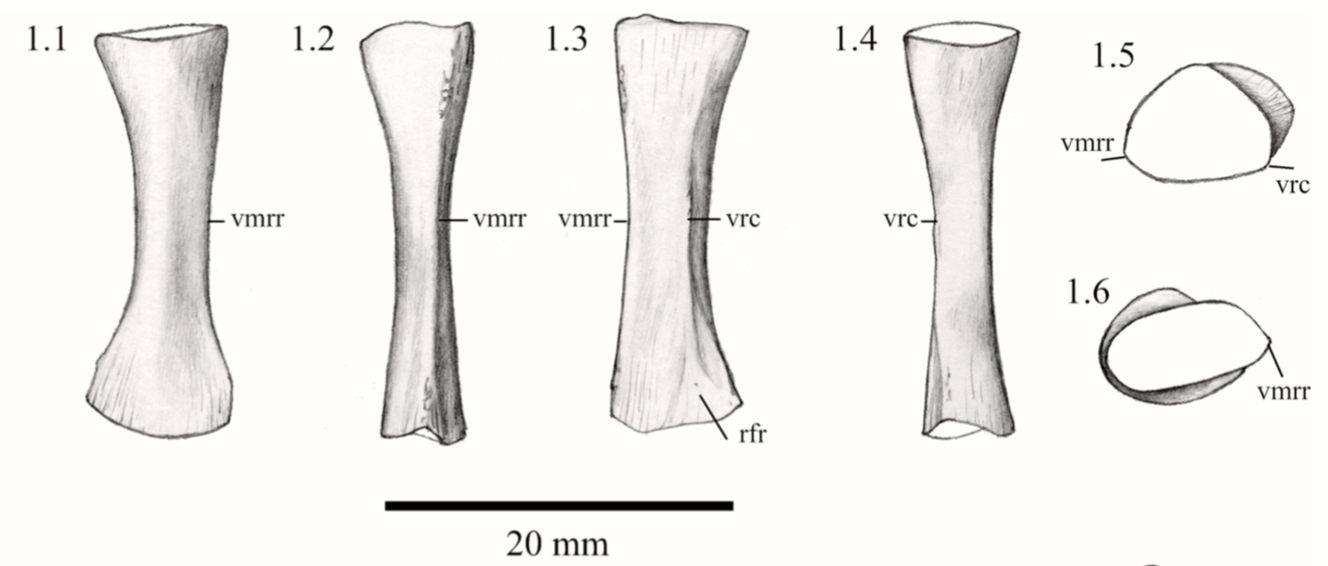
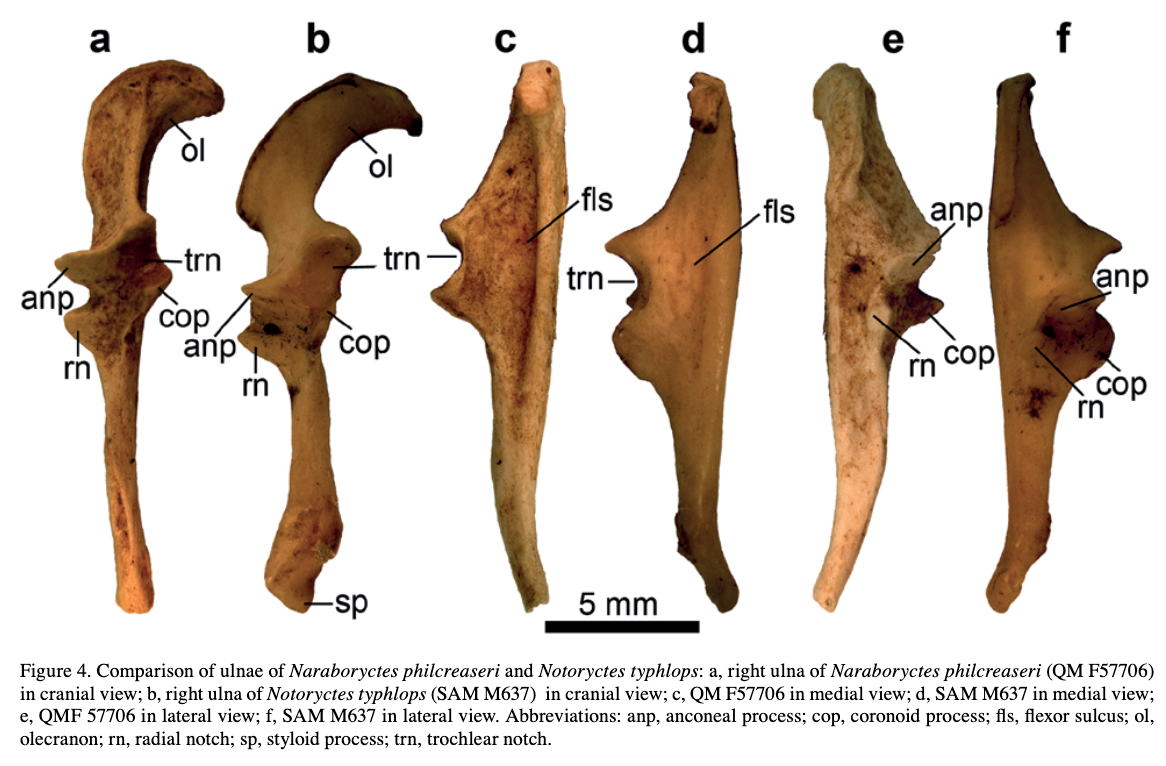
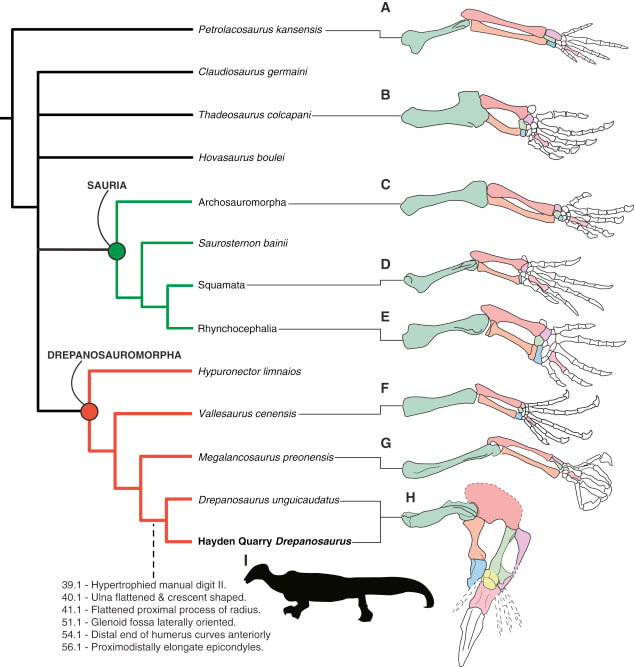
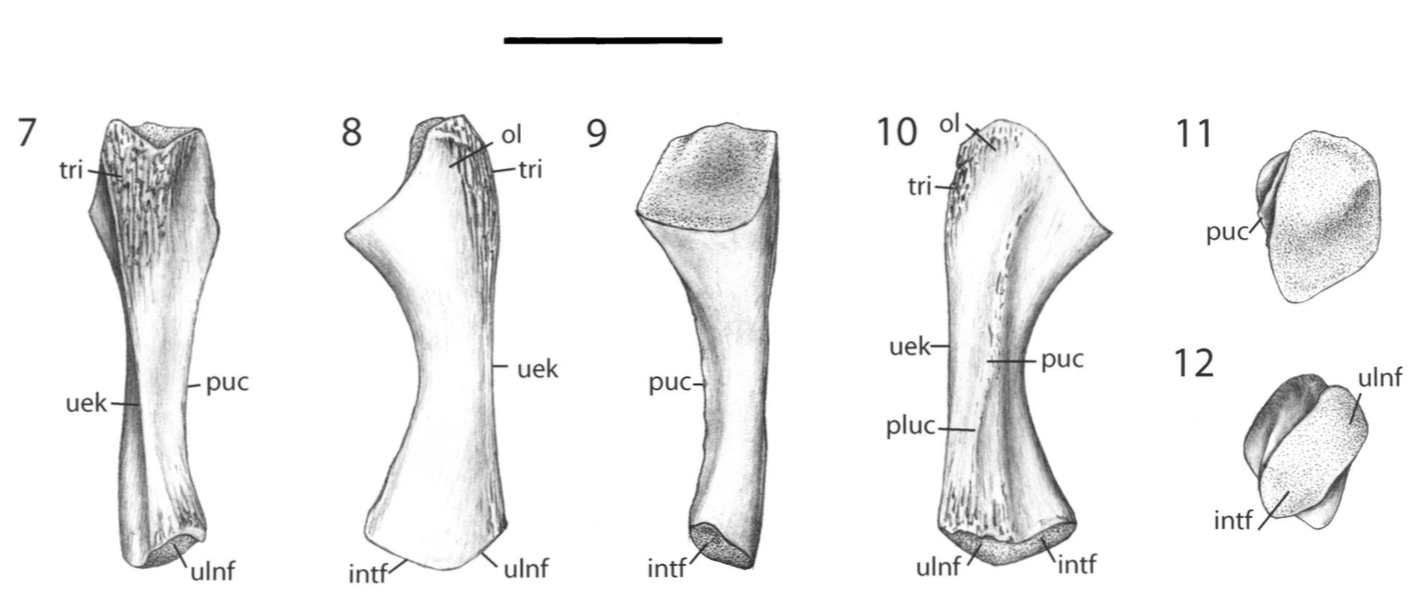
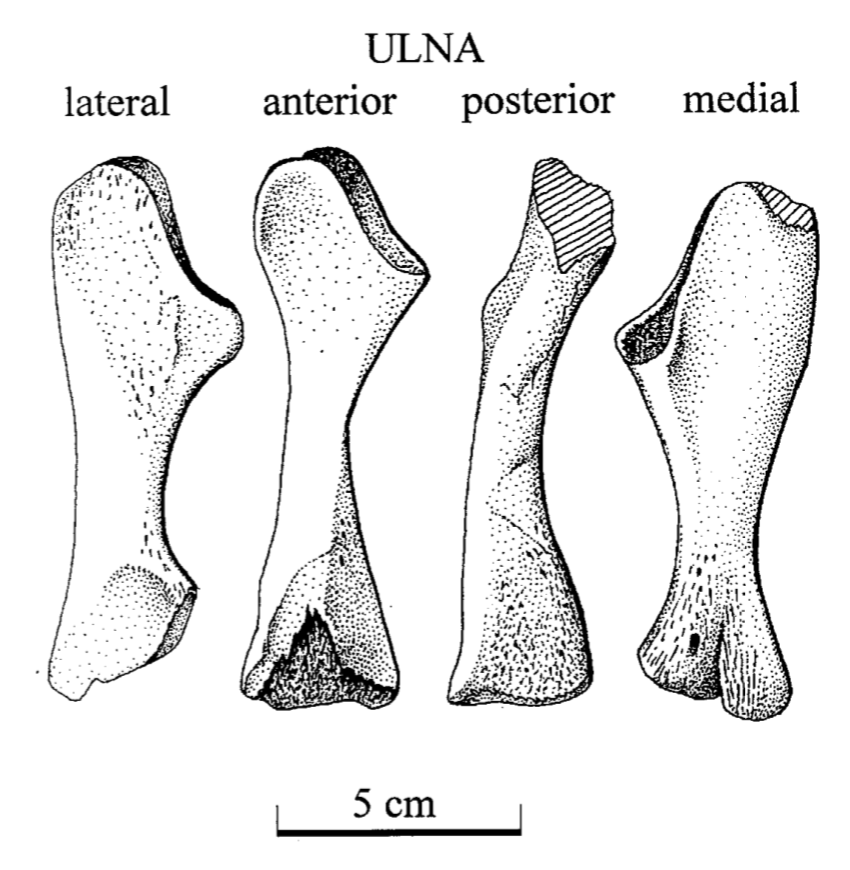
Unraveling the ulna The ulna is right next to the radius, but it is a little more distinctive and easier to identify. One of the main features that makes the ulna more recognizable is called the olecranon or the olecranon process (technically an olecranon is a process), and it's the bony protrusion that frames what we recognize as the elbow. The end of the olecranon, where it meets the humerus, has several major nerves, which is why hitting your "funny bone" evokes such a strong sensation. No word on whether temnospondyls also had a "funny bone" sensitivity. The olecranon is often very long and with a prominent facet in animals with very specialized lifestyles, especially those that use their forelimbs to burrow, like moles and echidnas. A similar morphology is often inferred to reflect forelimb-driven burrowing in extinct taxa, like the weird Triassic drepanosaurs, a group of reptiles that may also have had tree-dwelling species and a spike-like claw apparatus on the end of the tail.
The temnospondyl ulna is rather plain, which is probably not that surprising considering the radius. No taxon is known to have a markedly complex ulna that might suggest a specialized lifestyle. Today's amphibians are not big forelimb diggers; they often prefer to use their heads or their hindlimbs, and while they may scratch at dirt with their forelimbs, they are not expert burrowers in this fashion like moles. There are obvious differences in whether the olecranon is squared-off, with a slight cup-like excavation, or with a more pronounced socket, but because this also changes a lot throughout ontogeny, it has been hard to assign a particular functional import to these differences.
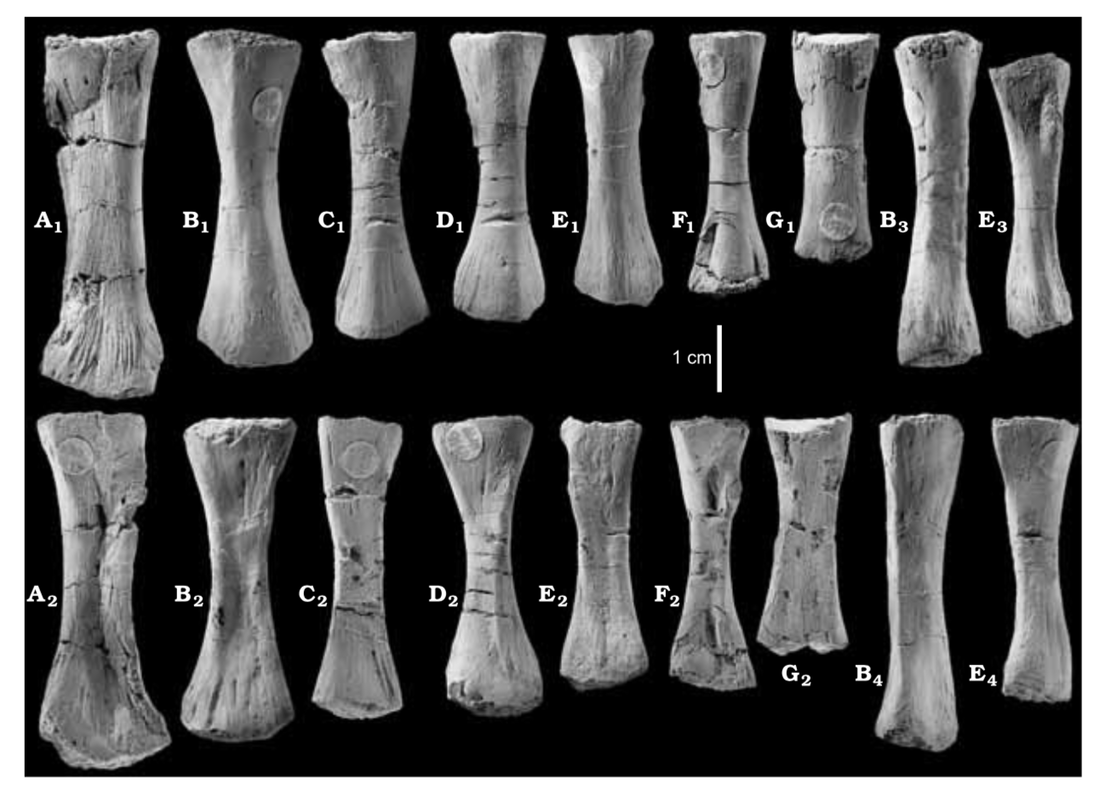
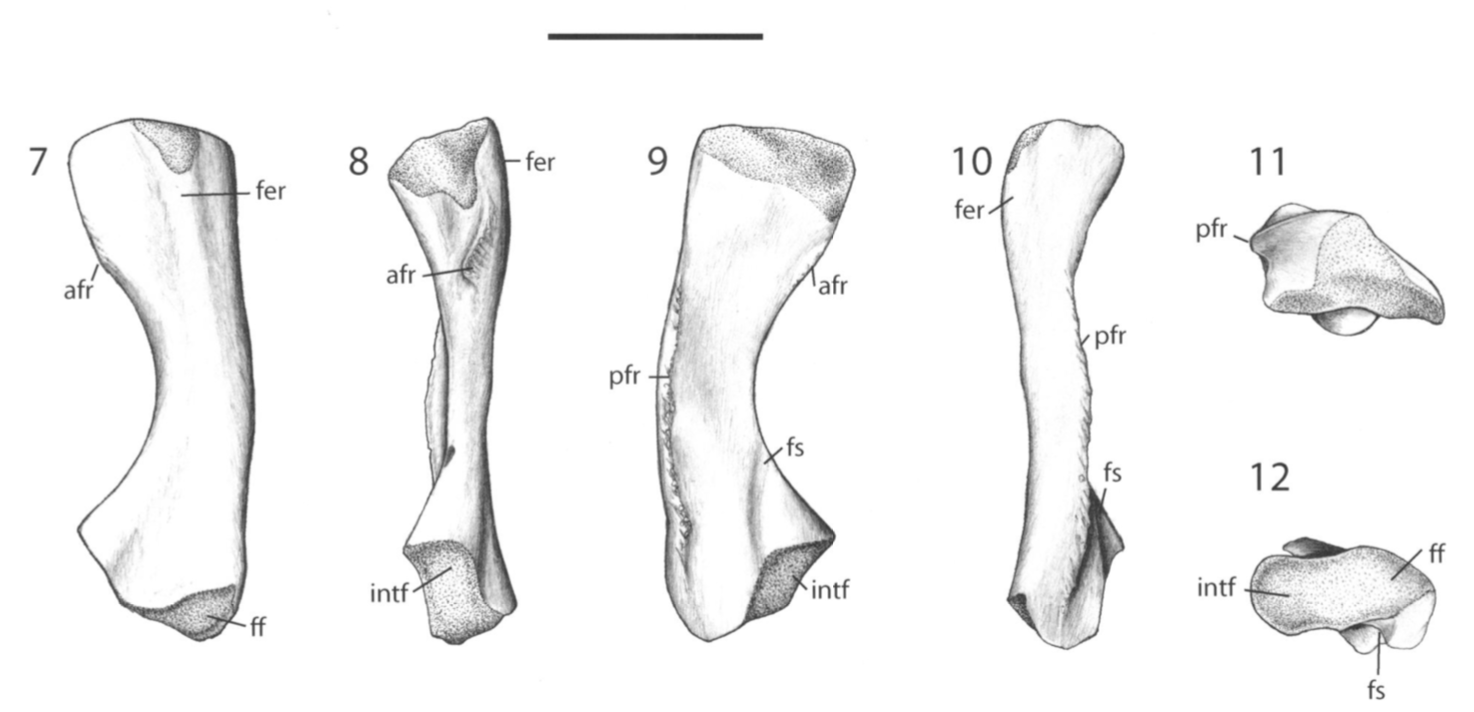
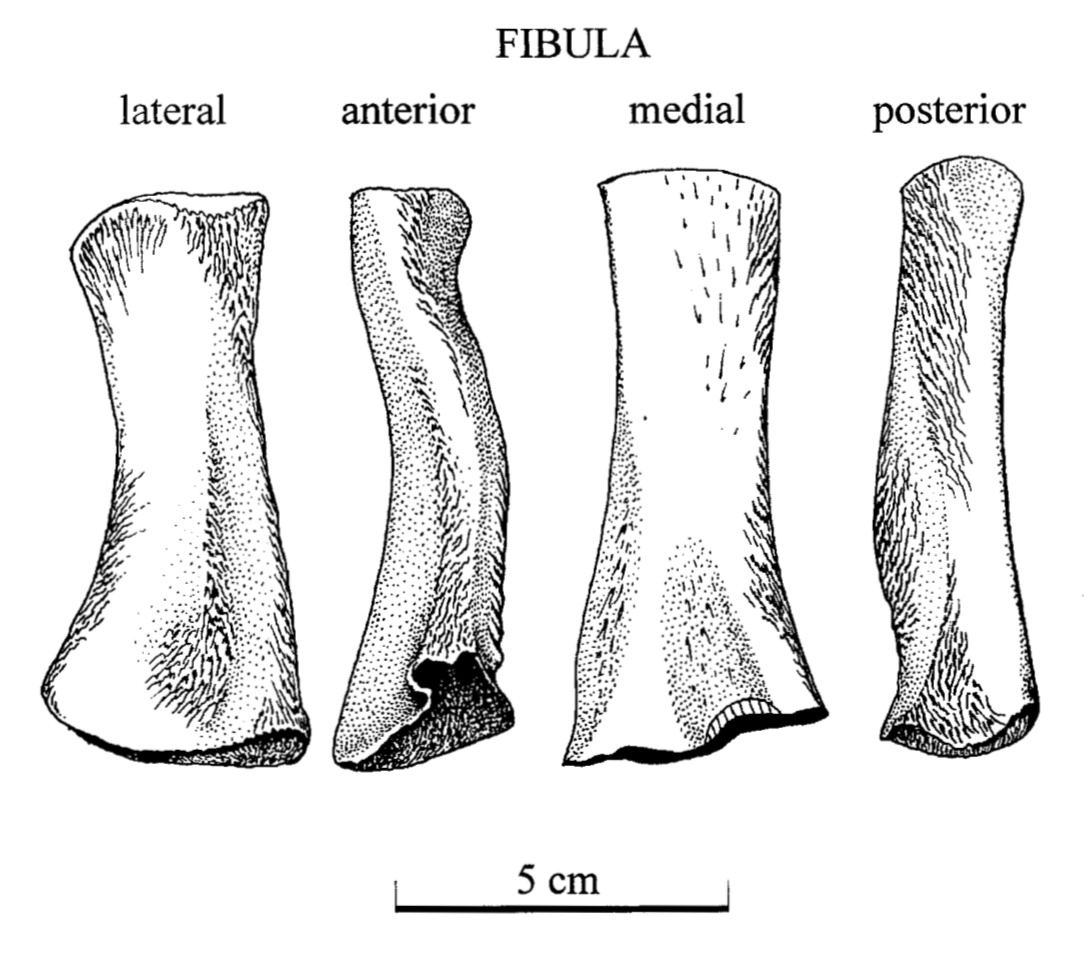
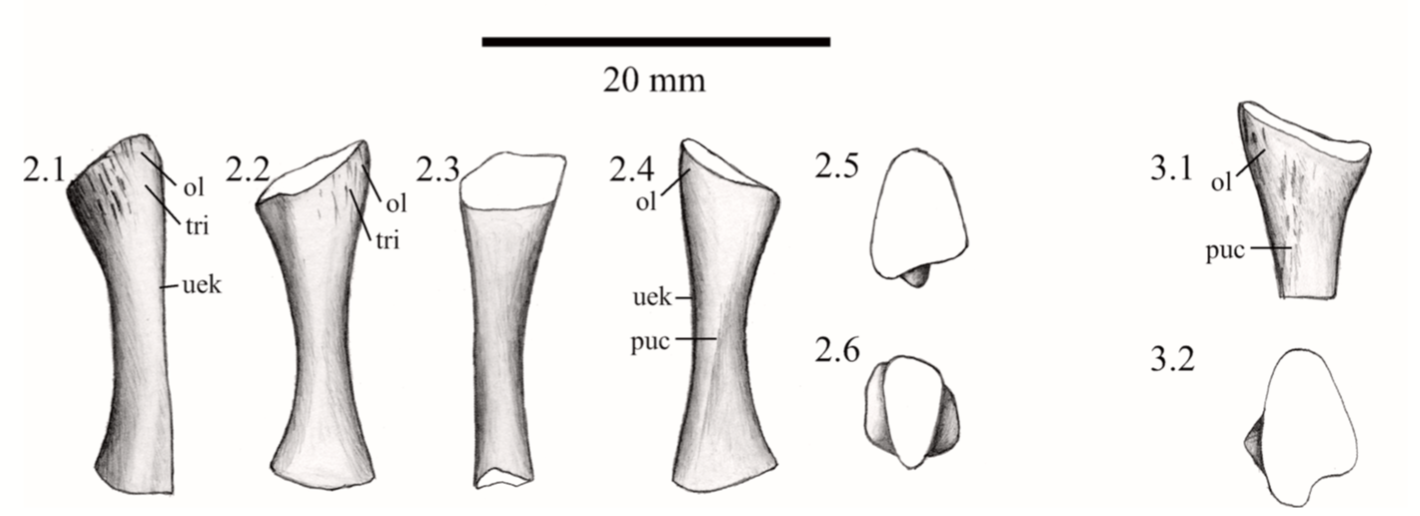 Ulna in multiple views from the Permian dvinosaur Trimerorhachis insignis (source: Pawley, 2007). Figuring the fibula The fibula is the radius of the hindlimb. Except it's usually a little curved and the entire bone tends to be flatter (oval cross-section), which makes it a little easier to identify in isolation. This has not really proven sufficient to figure out whether isolated fibulae can tell you much (or whether you can actually identify them as fibulae or to a particular taxon).
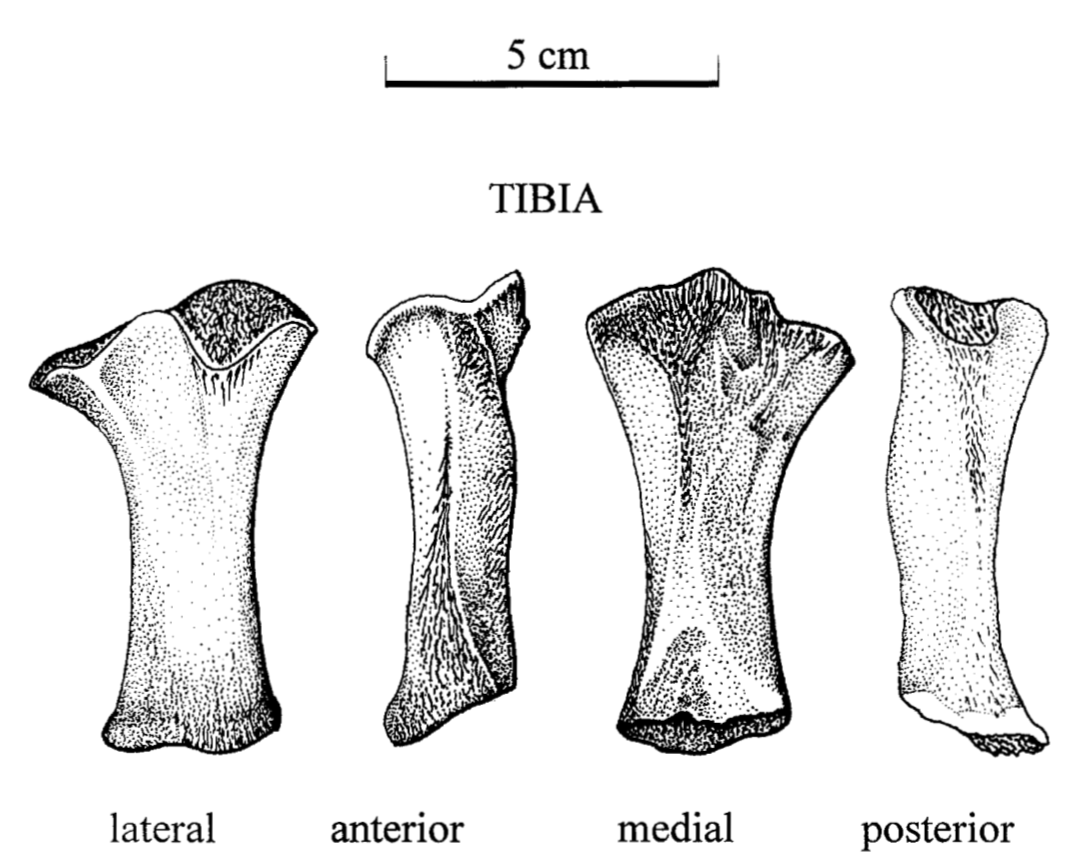
Tales of the tibia If the fibula is the radius of the hindlimb...well the tibia is the ulna of the hindlimb - a little more distinctive, definitely some differences, importance of said differences is not apparent. Unlike the ulna, however, the tibia is never markedly modified for a specialized strategy like digging because hindlimb-powered burrowing is much less elaborate than forelimb-powered burrowing (and also not as common). And thus while the knee is the elbow's equivalent (and why banging your knee also elicits the same expletive-loaded outburst), there is no long process or deep socket like we might see in the form of the olecranon on the ulna. The main aspect in which the tibia tends to differ is in the relative size of the two ends; disproportionate expansion of the proximal end (next to the knee) tends to be associated with inferred terrestrial capabilities.
A helping hand
Modern amphibians have not really done very much with their hands or feet other than develop soft tissue structures that we wouldn't likely find in the fossil record. This might include toepads for increased adhesion when climbing or webbing between the digits for gliding (yes, some frogs do glide). Mostly, amphibians just change the number of fingers or toes. A good example is a long-bodied aquatic salamander called Amphiuma, which lives in the water. There are different species of Amphiuma with one, two, or three toes. They are appropriately named the 'one-toed,' 'two-toed,' and 'three-toed' amphiuma. Most amphibians have four fingers and five toes, and usually, reduction in the number is often accompanied by more obvious changes that indicate that the limbs in their entirety are not being used as much for locomotion (as with the amphiuma).
Because we don't know much about the hands and feet, it's often hard to even identify isolated elements (having published on something with a foot myself, I know how confusing each bone can be when it's not in the exact same profile as whatever you're comparing to). We just don't know much about how much variation there is between species, or what might change throughout development. As a result, there is practically nothing that we can say about the ecology of temnospondyls from their hands or feet alone. Summary The short and skinny is that most temnospondyl limb elements are not actually that useful in isolation. Identifying something as a temnospondyl in general is pretty easy, but getting more specific than that, which is what we usually want, is not. Issues like ontogenetic immaturity, paedomorphosis (retaining juvenile features into adulthood), and intraspecific variation can all throw a wrench in certain qualitative features like "how well developed is this process?" Fortunately, we have some alternative means to tackle these questions on limbs from another angle. You may have gotten this far and thought, "hey, isn't there something about cutting limb bones to figure out how old they are?" There is (histology), and it can help us infer the lifestyle of various extinct tetrapods, but that's such a big topic that I'll cover it next week!
Comments are closed.
|
About the blogA blog on all things temnospondyl written by someone who spends too much time thinking about them. Covers all aspects of temnospondyl paleobiology and ongoing research (not just mine). Categories
All
Archives
January 2024
|





 RSS Feed
RSS Feed
