|
Title: Computed tomographic analysis of the cranium of the early Permian recumbirostran 'microsaur' Euryodus dalyae reveals new information of the braincase and mandible. Authors: B.M. Gee, J.J. Bevitt, R.R. Reisz Journal: Papers in Palaeontology DOI to paper: 10.1002/spp2.1304
Gymnarthrids 101First off, if you want a little more about 'microsaurs,' pop over to my summary of my PeerJ paper on Llistrofus from last year. We're back with more 'microsaur' magic, this time on the gymnarthrid Euryodus. In the longstanding debate over whether 'microsaurs' are in fact crown amniotes, not stem amniotes or stem lissamphibians, gymnarthrids are of great interest because they're among the most similar to eureptiles among the recumbirostrans (derived 'microsaurs' with recumbent snouts for digging). Specifically with respect to Richards Spur, material of Euryodus was long thought to be fairly common based on distinctive bulbous teeth until those teeth were found attached to the jaw of a captorhinid eureptile, Opisthodontosaurus (Reisz et al., 2015). Recumbirostrans are a weird batch, so gymnarthrids are sort of the "normal ones" - they don't have giant holes in their head like hapsidopareiids or lysorophians or extensively co-ossified braincases like brachystelechids or deeply vaulted skulls like ostodolepids. They're just sort of flat and uh...triangular. What gymnarthrids do have is that they're fairly common. A lot of other recumbirostrans are only known from one or two specimens, but there's a fair bit of Euryodus and its occasional doppelgänger, Cardiocephalus, floating around the early Permian of Texas and Oklahoma. We don't know too much about ontogeny in recumbirostrans, but the gymnarthrids are some of the few taxa where we have at least some info (Anderson & Reisz, 2011).
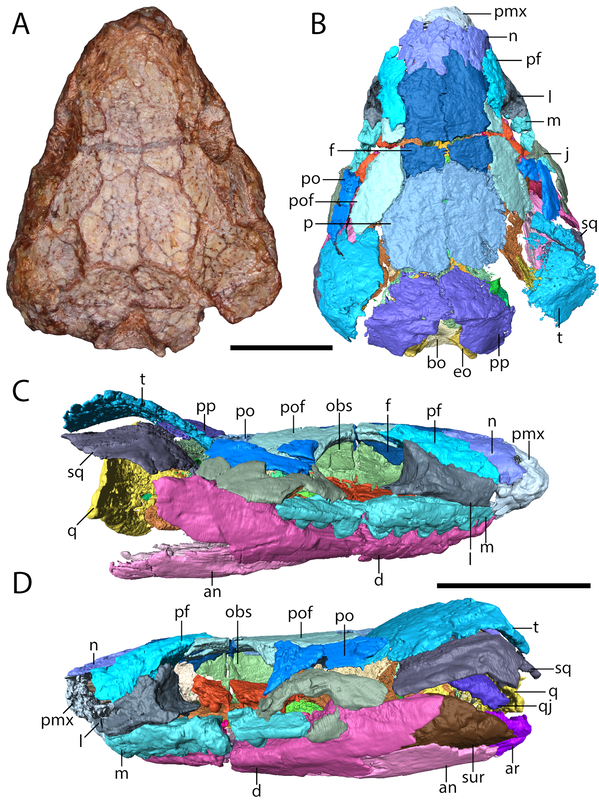
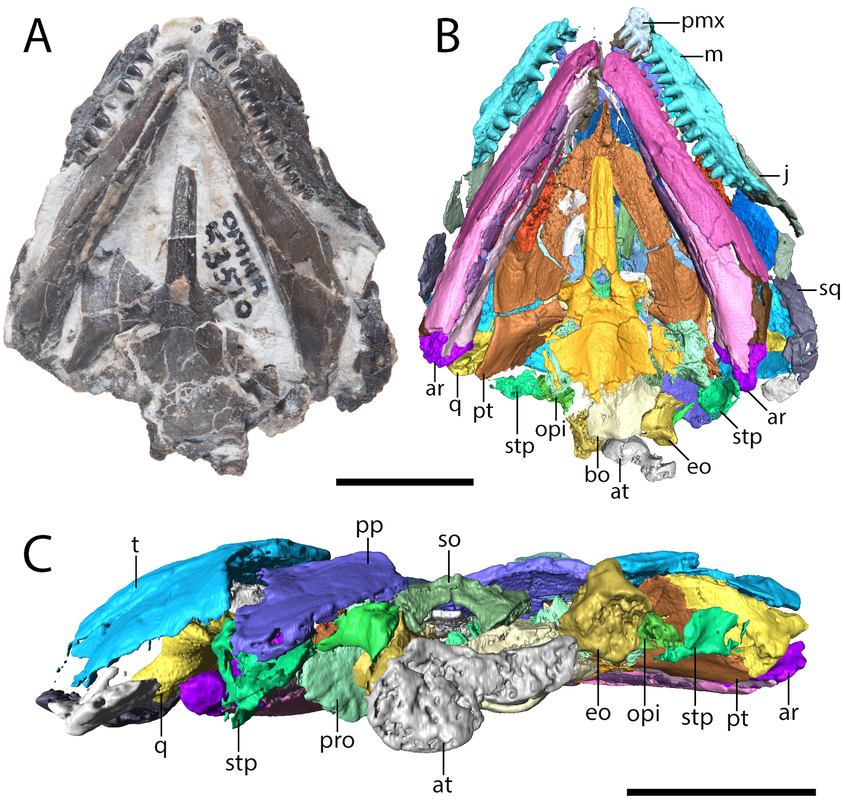
Skull of a referred specimen of Euryodus sp. from Richards Spur in...all of the views (figs. 7-8 from the paper). The many flavours of EuryodusThe Euryodus of this paper, Euryodus dalyae, isn't the OG Euryodus; that honour belongs to Euryodus primus from the early Permian of Texas, named in 1939 by American paleontologist Everett Olson. Another species of Euryodus, E. bonneri, was described from Kansas in 1981, but it was recently (well not too recently I suppose) demonstrated to be a different taxon, Proxilodon (Huttenlocker et al., 2013). Euryodus dalyae comes from Oklahoma, but not originally from Richards Spur; instead, it comes from the fairly notable South Grandfield locality. It was first described as Cardiocephalus cf. C. peabodyi, which speaks to the similarities between these genera (Daly, 1973), but it was subsequently named as a species of Euryodus after (Eleanor) Daly in 1978 by Bob Carroll and Eleanor Gaskill in 1978. Euryodus dalyae was reported from Richards Spur by MacDougall et al. (2017), which was apparently based on my advisor seeing the above specimen labeled as such in collections; there is no other published record.
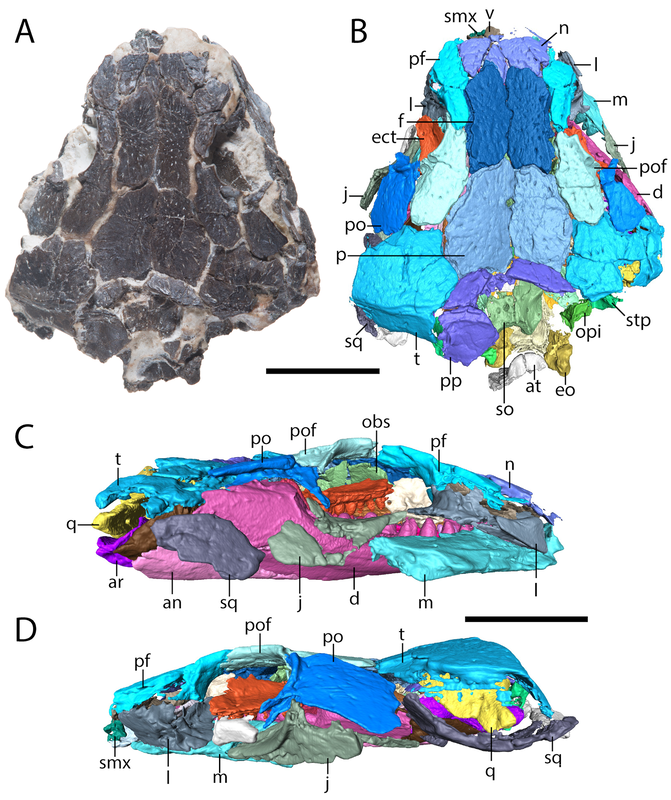
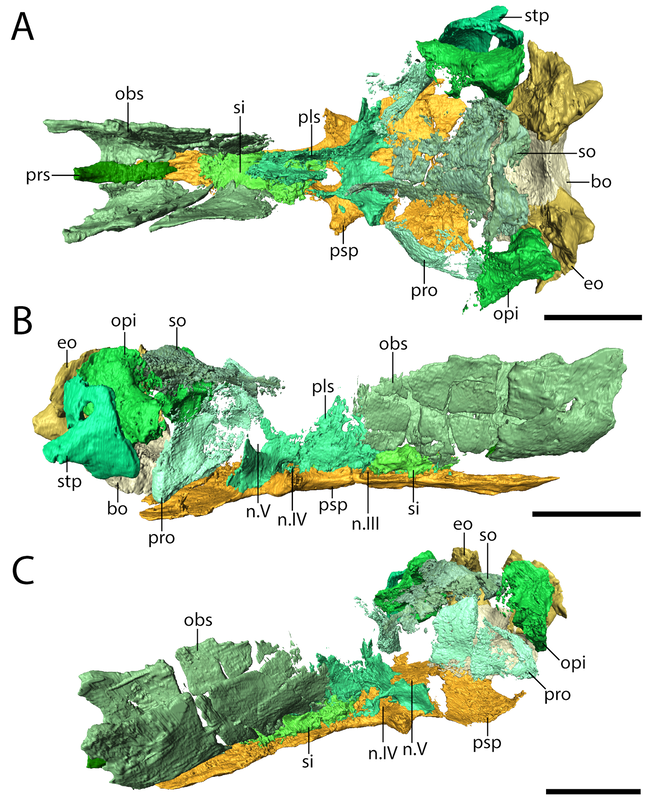
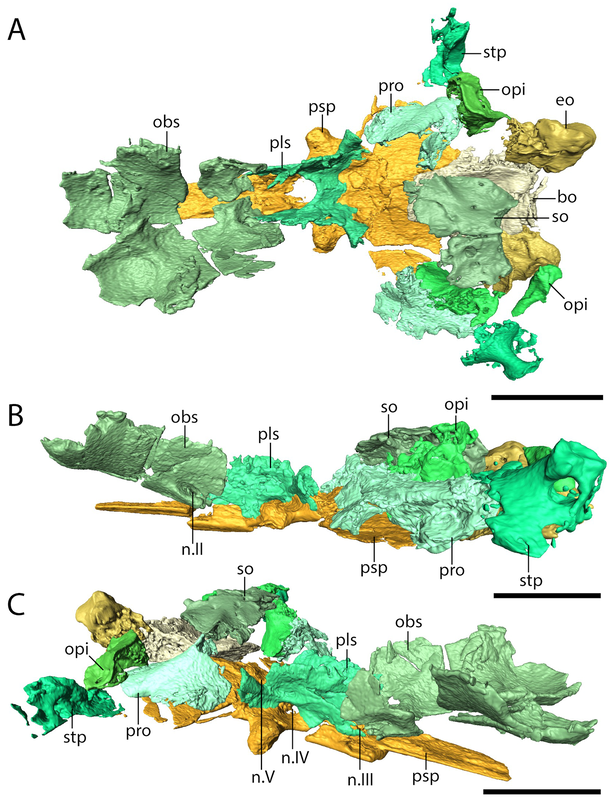
But this there actually Euryodus dalyae from Richards Spur? One of the major differences that we noted between the holotype from South Grandfield and the purported specimen from Richards Spur was in the braincase. The specimens aren't too differently sized, but the holotype (LEFT above) has a bunch of braincase ossifications found in other recumbirostrans but not in the RS specimen (RIGHT above). The two to note are most visible in part A of each (the dorsal view); the holotype has a presphenoid (prs) and an ossification of the subiculum infundibulum (si; try saying that five times fast). There's a few other differences that could be suggested to be ontogenetically influenced, but we just don't have enough information to say at present, and because the specimens are pretty similar in size, some of these transformations would seem to be very abrupt. Based on this, we elected to just refer the Richards Spur material (which includes a less complete skull that lives at the Field Museum) to Euryodus sp. Like the other Richards Spur 'microsaurs,' we don't get very many that fell into the fissures
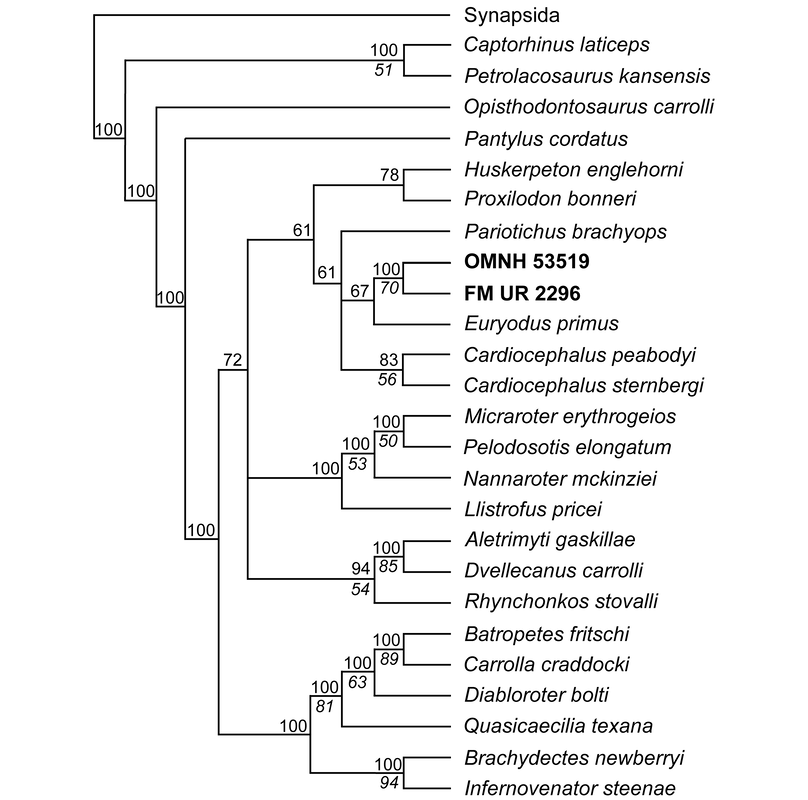
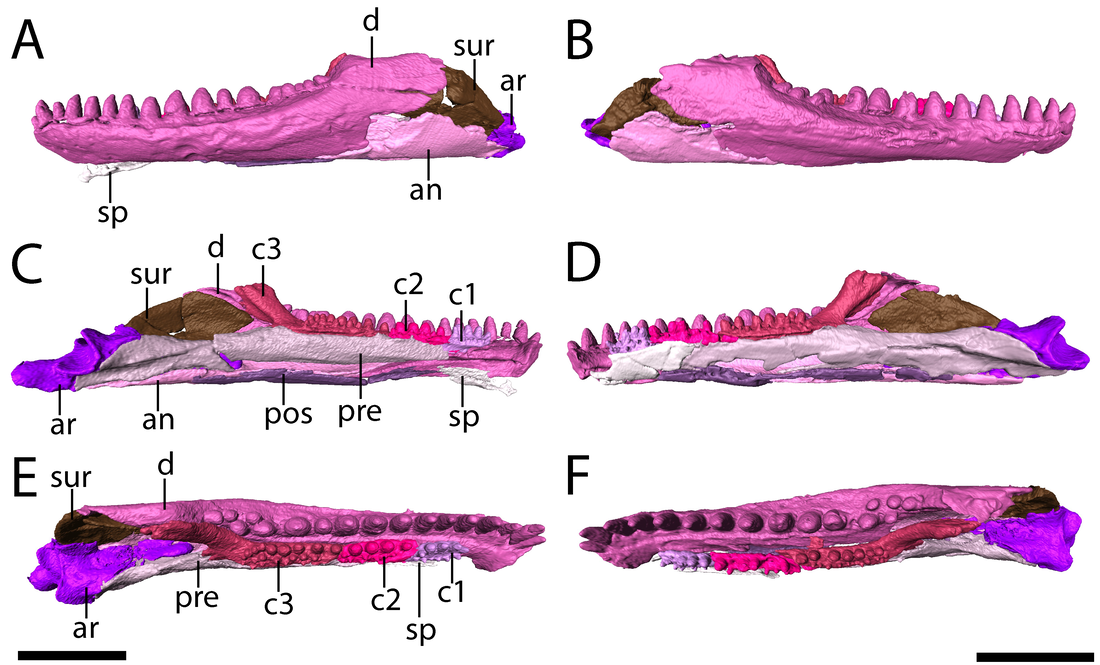 Lower jaws of the Richards Spur Euryodus. 'Microsaurs' have also been suggested to be early reptiles, an idea most recently revived by my friend Jason Pardo and colleagues (Pardo et al., 2017) who recovered recumbirostran 'microsaurs' as the sister group to captorhinids. This is based on a growing body of work identifying increased similarities between these clades, but the idea of 'microsaur'-captorhinid closeness is not new and has been debated for decades. I am a particular fan of Peter Vaughn's 1962 paper, cheekily titled "The Paleozoic microsaurs as close relatives of reptiles, again." There is of course, the alternative and similarly longstanding hypothesis that 'microsaurs' are actually closely related to modern amphibians, a stark contrast to their possible reptile affinities. One can identify homologous features (or analogous depending on your take) across the skeleton of 'microsaurs' and captorhinids, but one area that hadn't turned up anything yet was tooth rows...until now! As you can see below (parts E and F), this 'microsaur' had a partial second tooth row on the main element of the mandible, the dentary ('d'). One could argue that the teeth situated more internal on the coronoids ('c1,' 'c2,' and 'c3') add yet more mores to form a continuous toothy surface. We'd known about the extensive palatal dentition of some 'microsaurs,' but nobody's ever identified a second marginal tooth row until now. This row is extremely hard to ID when the jaws are articulated with the skull, and we could only clearly parse it out in the digital data. In captorhinids, multiple tooth rows aren't the product of absurd tooth growth, but the retention of several generations of teeth due to disproportionate jaw remodeling. 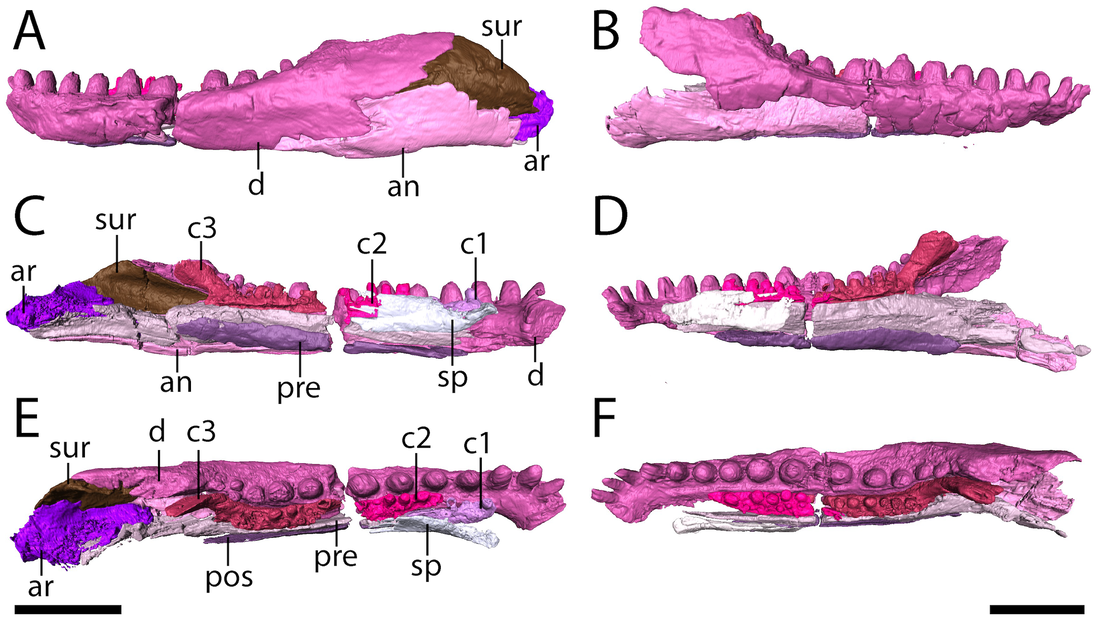 Lower jaws of the holotype of Euryodus dalyae. Refs
David Marjanović
3/15/2020 02:01:34 pm
"There is of course, the alternative and similarly longstanding hypothesis that 'microsaurs' are actually closely related to modern amphibians, a stark contrast to their possible reptile affinities." Comments are closed.
|
About the blogA blog on all things temnospondyl written by someone who spends too much time thinking about them. Covers all aspects of temnospondyl paleobiology and ongoing research (not just mine). Categories
All
Archives
January 2024
|

 RSS Feed
RSS Feed
