|
Title: Revision of the Late Triassic metoposaurid “Metoposaurus” bakeri (Amphibia: Temnospondyli) from Texas, USA and a phylogenetic analysis of the Metoposauridae Authors: B.M. Gee; A.M. Kufner Journal: PeerJ DOI: 10.7717/peerj.14065 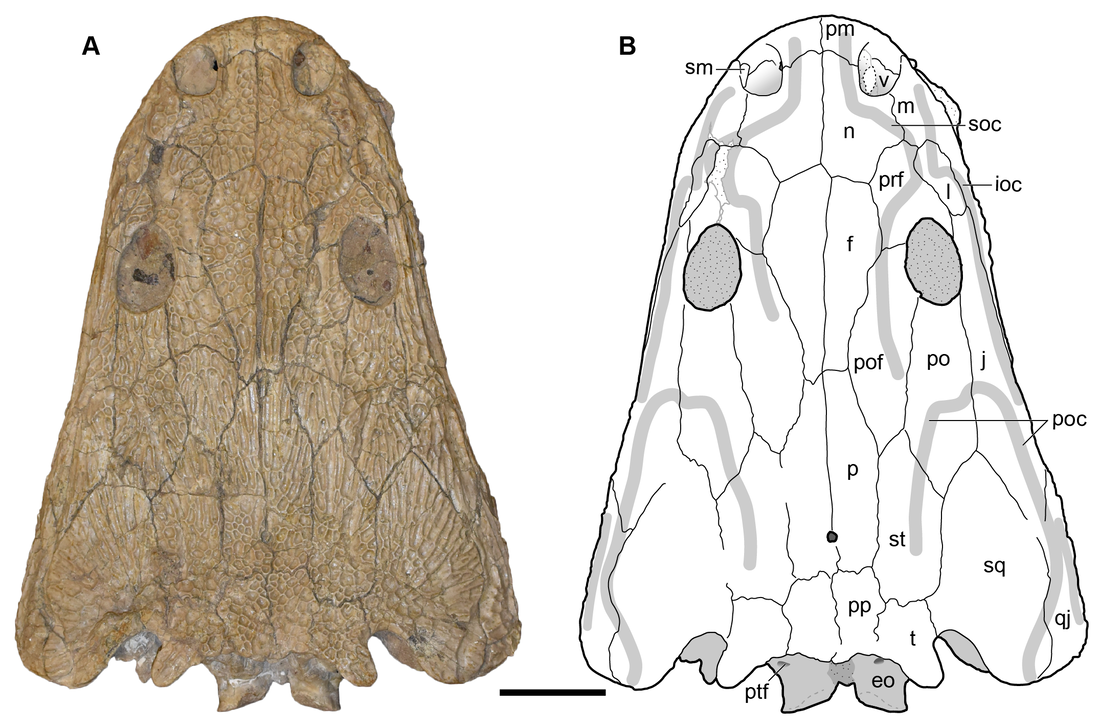 Photograph and interpretive line drawing of UMMP 13820, a referred specimen of Buettnererpeton bakeri (Fig. 9 from the paper). General summary: Frequent readers of this blog are of course familiar with my love for metoposaurids, one of the most iconic groups of North American temnospondyls. It is no secret that metoposaurids were my "gateway drug" to the world of terrifyingly large and unrecognizable amphibians, and so they always have a special place in my heart. My previous research has largely focused on two of the three species, Anaschisma browni (ex. Koskinonodon perfectus, ex. Buettneria perfecta) and Apachesaurus gregorii, which are the two most common metoposaurids that we get in the Late Triassic of North America. If you go to a museum and see a metoposaurid (most major museums in the U.S. have a metoposaurid on display), it's likely Anaschisma browni, although the label may be two or three junior synonyms out of date. The third species, known only from two sites in Texas and one site in Nova Scotia, doesn't get as much press - it was last (re)described in 1932! Ninety years later, my buddy Aaron Kufner and I are pleased to produce a full redescription of this taxon, long referred to as 'Metoposaurus' bakeri, based on a reexamination of material from the type locality that now lives at the University of Michigan Museum of Paleontology (UMMP) collections in Ann Arbor. With a thorough redescription spanning 134 print pages (probably too thorough, even by my own standards), we provide extensive documentation of the skeletal anatomy of this species (1930s-era drawings have their limitations), conducted more phylogenetics analyses to test the relationships of metoposaurids (this has gotten no better since the 1930s), and ultimately concluded that this species cannot be placed in an existing genus like Metoposaurus (otherwise only known from Europe). To that end, we created a new genus name, the mouthful Buettnererpeton, which honors a longtime fossil preparator at the UMMP, William H. Buettner, who worked extensively with E.C. Case, the museum curator who named the species in 1931. The suffix comes from -herpeton, meaning 'creeping animal' in Greek, which is a common component of names of early reptiles and amphibians. Our taxonomic act has implications for biostratigraphy (relating distantly situated rocks based on which taxa occur in them), both globally and within North America, and we discuss everything from future work needed on metoposaurids to why their phylogenetic relationships are so badly resolved. This is a 'boundary-crossing' project that originated when I was a Ph.D. student in the summer of 2019 and has only now made it to the finish line, so it is particularly memorable for me in that regard. Entangled in taxonomy
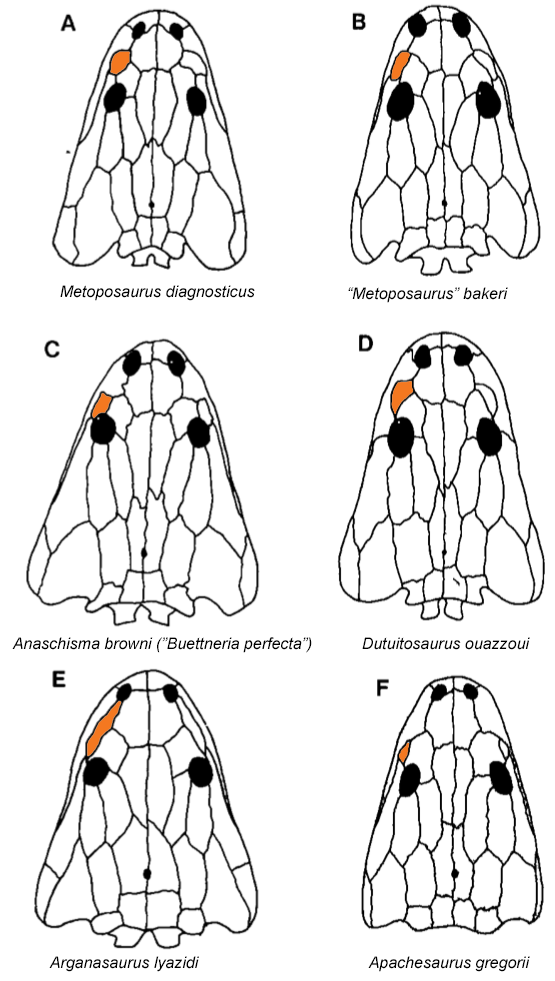
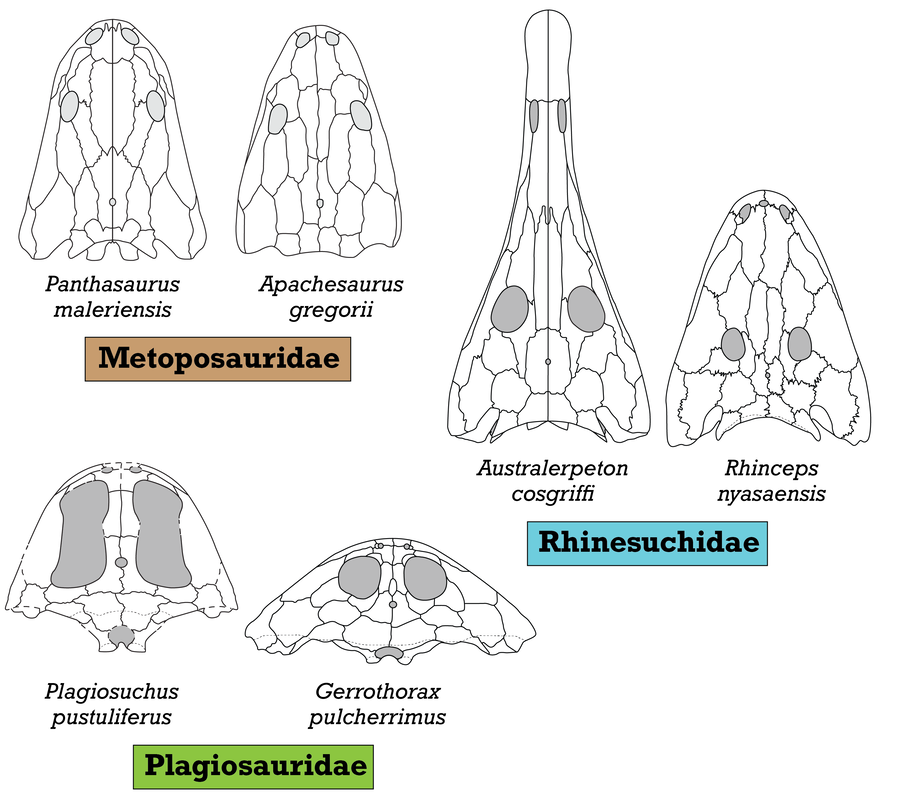 Comparison of semi-arbitrarily picked "end-members" of the morphological range for three temnospondyl families: Metoposauridae, Rhinesuchidae, and Plagiosauridae. Silhouettes adapted from Chakravorti & Sengupta (2018), Spielmann & Lucas (2012), Eltink et al. (2016); Schoch & Milner (2000); Damiani et al. (2009), and Schoch & Witzmann (2012). Most of the excessive taxonomic splitting (naming new species based on features that are not considered reliable for taxonomic differentiation, like size of the type specimens), has been historically concentrated in North America and Europe, but it's been fairly stable in North America for a few decades now, with everyone agreeing that there are three valid species. I've tackled two of these in recent years, Anaschisma browni (the one that keeps getting renamed, much to the chagrin of museum exhibit designers) and Apachesaurus gregorii, but the remaining metoposaurid is the least well-studied and thus the most taxonomically ambiguous: "Metoposaurus" bakeri (which you can find with pretty much every combination of genus + species, both with an without quotes, e.g., Koskinonodon bakeri, Buettneria bakeri). 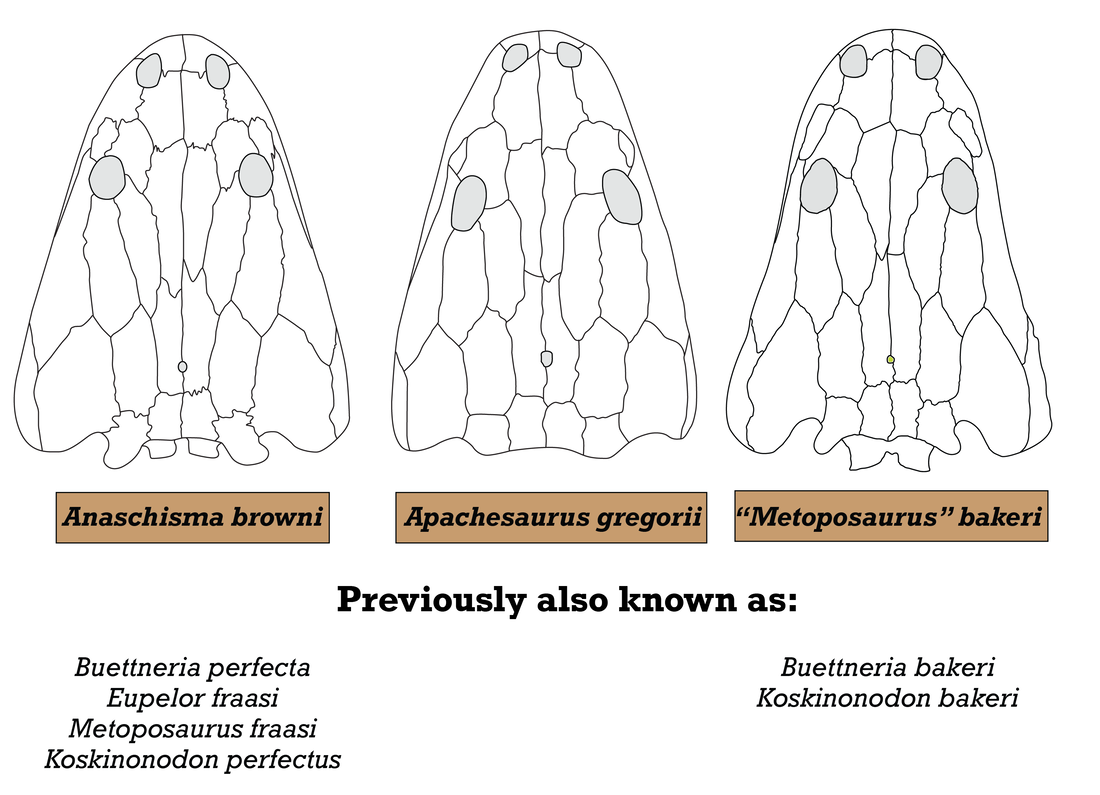 Comparison of the morphology and taxonomic history of the three recognized North American metoposaurids. Silhouettes after Case (1922); Spielmann & Lucas (2012); and this study.
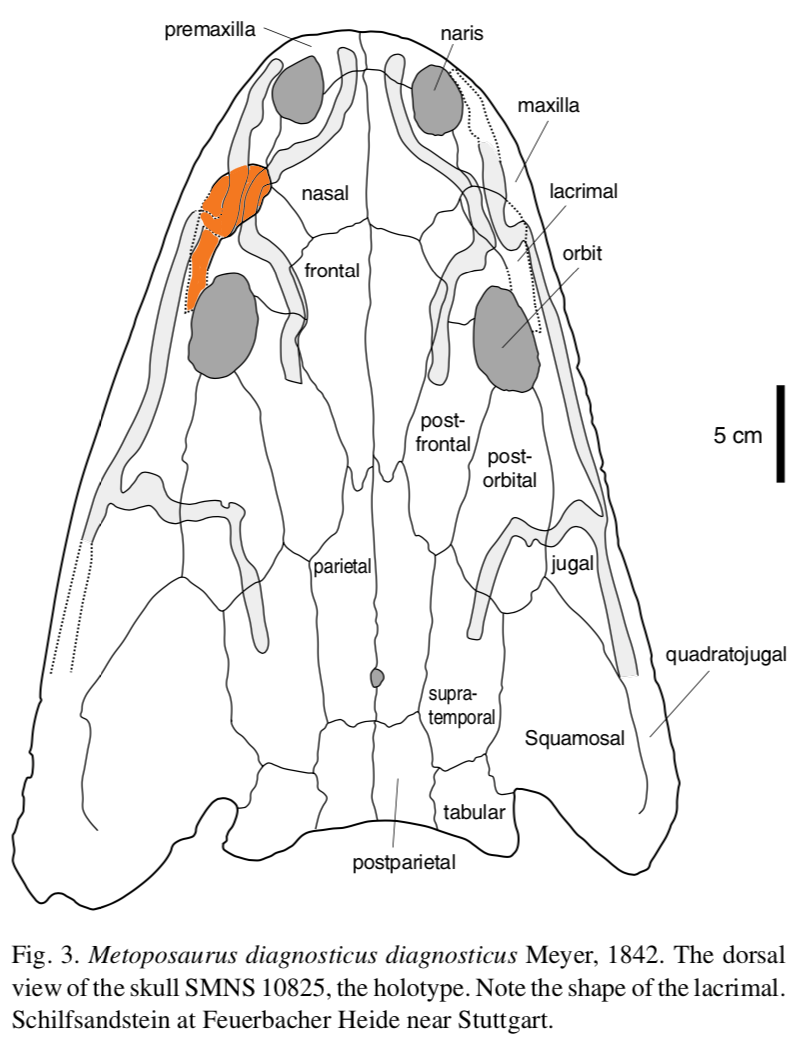
The rationale for the updated taxonomy is entirely normal for paleontology but illustrates the subjectivity associated with how scientists differentiate at a given taxonomic scale. "Metoposaurus" bakeri was primarily allied with Metoposaurus diagnosticus because both taxa purportedly shared the condition of having a lacrimal bone that was separated from the orbit, in contrast to Anaschisma browni (then "Buettneria perfecta"), which has a lacrimal that enters the orbit (see above on right). That's it, one binary feature. "Metoposaurus" bakeri was then differentiated from M. diagnosticus by several qualitative features, like a proportionately longer lacrimal and a smaller area of radiating grooves on the clavicle. Both are (1) really continuous, not discrete, and (2) shared with A. browni. Other shared features have also been argued to separate the North American taxa from Metoposaurus proper from Europe, like the relative size of circular pitting on the interclavicle (see below). All of this is to say that an argument could have been made to unite "M." bakeri in the same genus with A. browni, and to use the lacrimal-orbit relationship as their one differentiating feature. 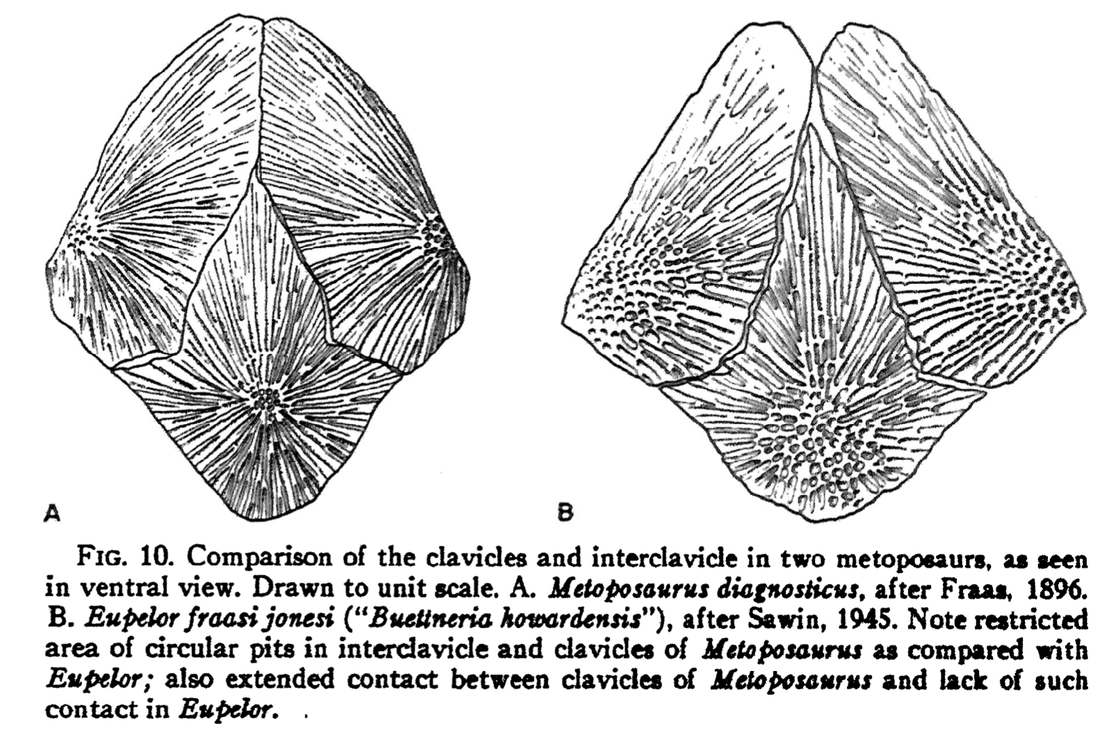 Illustrations of the pectoral girdle of the European (A) and North American (B) metoposaurids, with larger areas of circular pitting on the underside of both the clavicle and the interclavicle in the N. American taxa (B). Figure from Colbert & Imbrie (1956).
Breaking the binary This convoluted history brings us to this study. Whether "Metoposaurus" bakeri really belongs in Metoposaurus or not has big implications for both metoposaurid taxonomy and biostratigraphy. If "M." bakeri was placed in Metoposaurus, this would further undermine the historical use of the lacrimal-orbit relationship as a differentiator of metoposaurids at the genus level. The reason why this is so problematic is not the undoing of precedent (a lot of historical precedent should be undone) but rather that the only reliable diagnostic cranial feature of Metoposaurus (in the sense of Brusatte et al., 2015, in which only the three European species belong to this genus) and Anaschisma is the lacrimal-orbit relationship. In other words, if "M." bakeri were placed in either genus, it would mean that a lacrimal entering the orbit would no longer be diagnostic of that given genus, and therein, that would mean that many isolated skulls from North America or Europe would essentially be unable to be referred to a particular genus, let alone to a species. This would, for many reasons, be very bad for all paleontologists, not just academics. (It also illustrates that perhaps Anaschisma should be synonymized with Metoposaurus, as it was in the 60s and 70s). 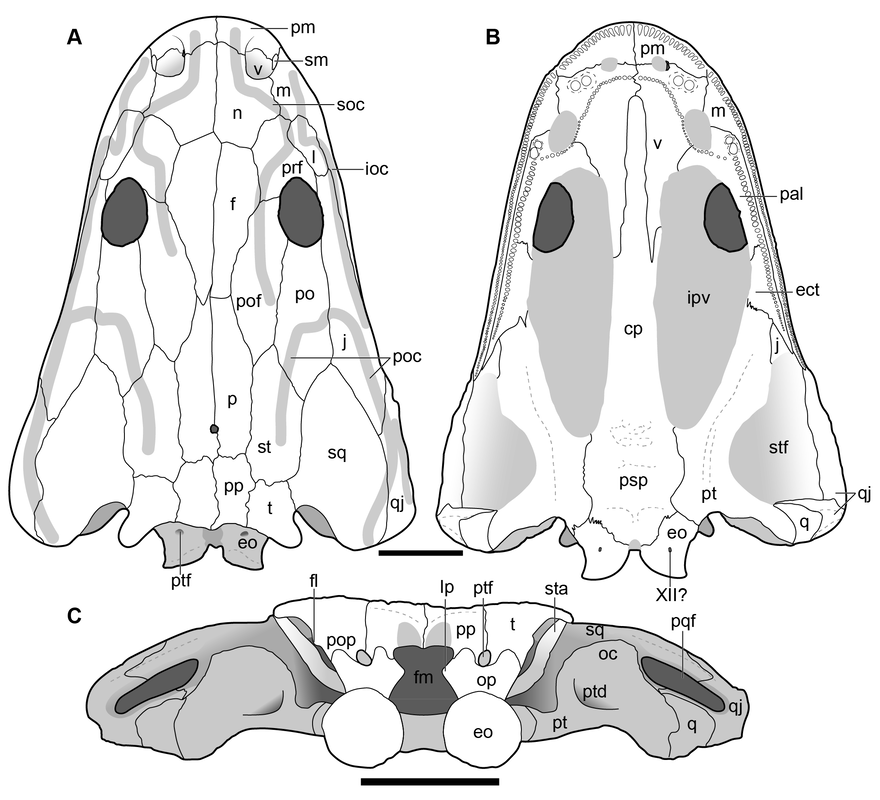 Our new cranial reconstruction (full credit to Aaron for this one; Figure 5 in the paper). Scale bar is 5 cm long. However, there are options beyond trying to squeeze "Metoposaurus" bakeri into one of these two existing genera. Although I tend to be more of a 'lumper' than a 'splitter' in casual academic parlance (I tend to prefer synonymizing taxa than to create new ones), there was really no other option given the difficulty that we already have with differentiating metoposaurid genera. Hence, Buettnererpeton was born! This admittedly very wordy name takes its inspiration from the now-defunct Buettneria, a name created in 1920 by E. C. Case to honor a longtime colleague and fossil preparator at the University of Michigan, William Buettner. Finding some way to restore Case's homage was something that the collections manager, Adam Rountrey, had suggested to us back when we first visited in 2019, and combining Buettner's name with -herpeton, a common suffix for Paleozoic tetrapods (meaning 'creeping thing' or some variation of that), was the obvious choice. Note: for most of the study, we proceeded to write a graphically long anatomical redescription of Buettnererpeton bakeri, to the tune of 62 anatomical figures and 78 PDF pages of description. I will not bore the reader with most of the relevant details. They are very graphic. I somewhat resent myself for how much attention to detail we paid. The rest of this blog will keep going with more fun topics.
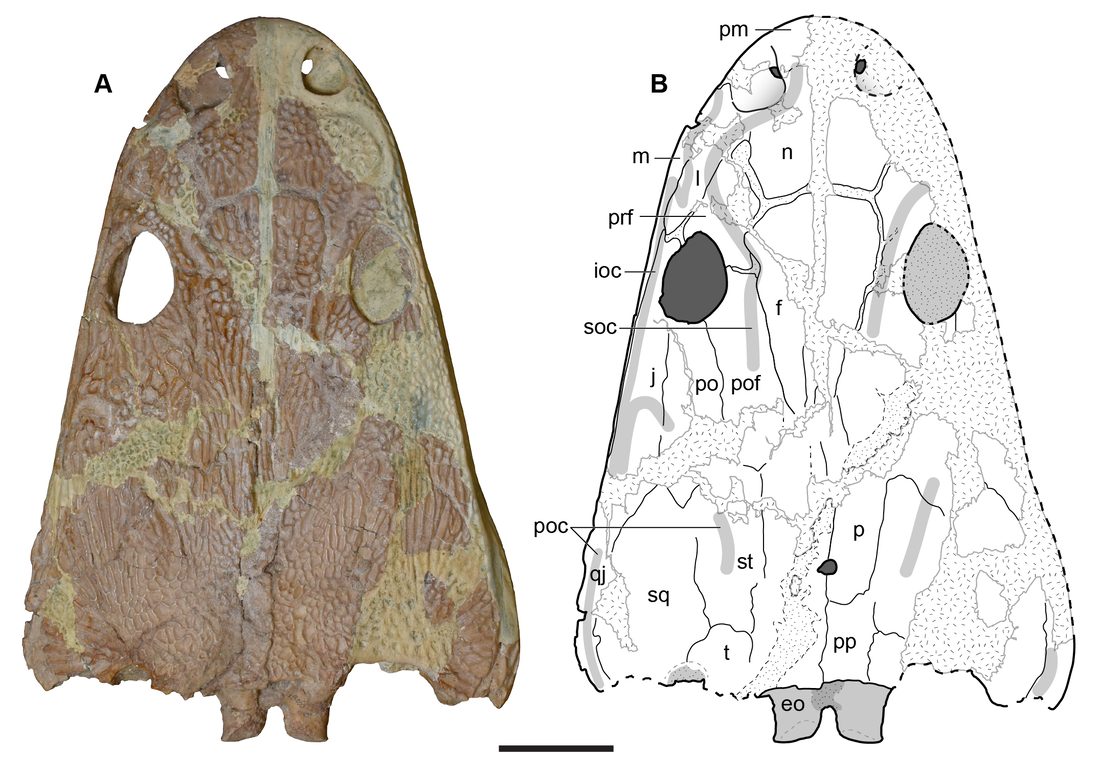
Photograph and interpretive line drawing of the dorsal surface of the holotype skull (UMMP 13055) of Buettnererpeton bakeri.
Perhaps I took a wrong turn One of the prominent regional patterns in the distribution of metoposaurids within North America is that there are very few on the eastern half of the continent, owing in large part of the much more limited exposures of Late Triassic rocks. Essentially all of the ones on display are from Arizona, New Mexico, or Texas (even the ones housed in east coast museums like the Smithsonian, American Museum of Natural History, and the Museum of Comparative Zoology). Most metoposaurid material from the eastern seaboard was collected during the construction of tunnels, which as you might imagine, was less than conducive for the collection of delicate fossils. As a result, practically none of it is diagnostic to a certain species, and it requires a fair bit of circular logic to even assert metoposaurid identity at all.  A photograph (graciously provided by Dr. Hans-Dieter Sues of the Smithsonian) and interpretive line drawing of YPM VPPU 021742. The one exception to this is a very nice natural mold of Buettnererpeton bakeri from Nova Scotia, also one of the few nice Mesozoic temnospondyl fossils from Canada. This specimen was first collected several decades ago (and now lives at Yale) and has long been the only occurrence of B. bakeri from outside of Texas. Although its identity hasn't been particularly controversial, Hans kindly gave us a good photograph, from which we reproduced an interpretive line drawing (not previously done), to corroborate its conspecificity with the Texas material. Its isn't that surprising that metoposaurids would have lived there - they are found in Morocco and western Europe, which would have been much closer to the eastern seaboard in the Late Triassic - but it does demonstrate the geographic range of one species. Really broad geographic ranges are only something that we observe in North America, likely due to a combination of the large size of North America compared to Europe, the heterogenous geography of the Late Triassic, and the heterogenous distribution of Late Triassic exposures today. But it's nonetheless important for indicating that metoposaurids were likely distributed across much of North America and are only unknown from many parts because of lack of Late Triassic exposures. Quite possibly, they were as widely distributed as common living fauna like raccoons. To each their own One of the longstanding practices in paleontology was naming new species when something was found in a new region. Of course, perceptions of geographic distance can be greatly exaggerated by the modern configuration, when in fact what are now disparate regions could have been closely situated in the geologic past. The general amalgamation of the continents into Pangea was largely consistent throughout the Triassic, as seen below. 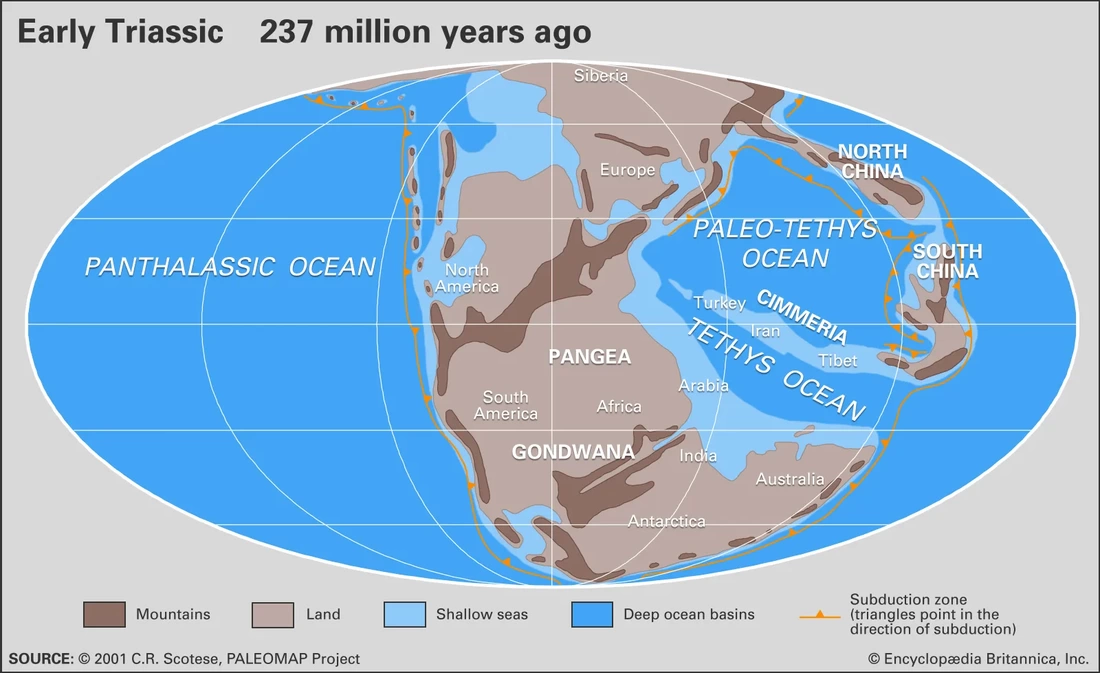 As a result, most of the metoposaurid-bearing regions are actually quite close to each other. Nova Scotia was very close to the Argana Basin of Morocco, from which many metoposaurids have been collected, closer in fact than it was to the southwestern United States. At the time, Nova Scotia was about as close to Germany as it was (is) to Arizona. The below figure from Brusatte et al. (2015) illustrates this well, demonstrating that there was a strong latitudinal constraint on the distribution of metoposaurids, which have yet to turn up in the well-explored Late Triassic of South America. 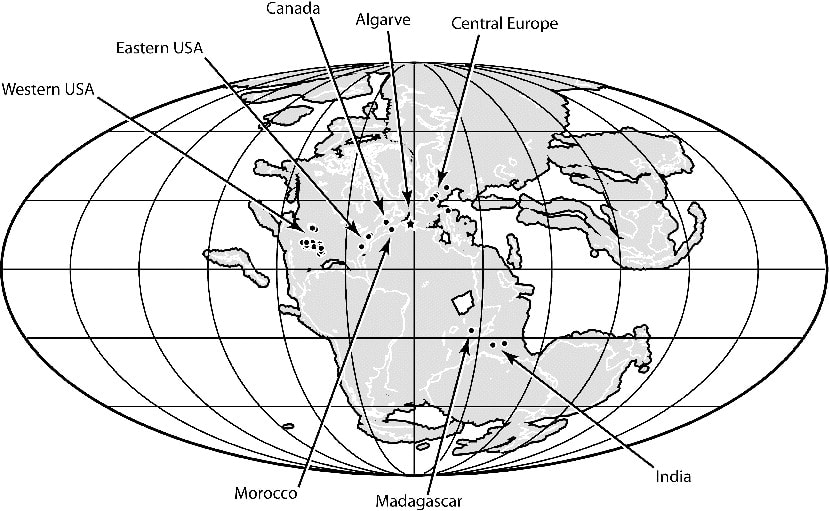 "Metoposaurus" bakeri figured somewhat prominently in the attempts of some workers to make correlations in relative age between geographically disparate regions around the world. Dating back to the old framework of Hunt (1993) in which Metoposaurus was found in both North America (viz. "M." bakeri) and Germany (viz. M. diagnosticus), certain workers honed in on Metoposaurus as a possible index taxon, a species or genus that is sufficiently common and widespread to serve as one of the markers used to correlate rocks deposited in disparate regions. At the time, no other genus and no species was found on more than one continent; most are actually quite restricted. However, old-school taxonomy is often conceptually suspect, and as a result, a lot of these index taxa have been diminished in biostratigraphic value. Sometimes it's shown that the occurrence in one region is not actually that taxon, so there is no direct correlation. Other times, original identifications were somewhat circular in logic or based on very fragmentary material, leaving a lot of doubt. This is not exclusive to metoposaurids, which has led to a stark bifurcation in utilized taxonomic frameworks of all sorts of Triassic tetrapods, with proponents of an ability to use terrestrial tetrapods for global biostratigraphy preferring older (outdated) frameworks that maintain the utility of this global system. This is likely an unstated rationale behind certain workers preference to maintain "M." bakeri in Metoposaurus, even after studies demonstrating that they do not actually share the lacrimal excluded from the orbit. Our formalization of a new genus for "M." bakeri puts the final nail in the coffin for the use of metoposaurids for global biostratigraphy (although I'm sure that proponents of this scheme will continue to push the old framework). With this, no metoposaurid genus or species is definitively found on more than one continent, and most are restricted to a single depositional basin. Although metoposaurids were clearly widely spread globally thanks to the formation of Pangea, it's not surprising that taxonomic divisions along some of the modern continental boundaries as the supercontinent started to break up. Metoposaurids have never been shown to have saltwater tolerance or to be particularly capable of much terrestrial locomotion (i.e. they were limited to dispersal in freshwater). Consequently, even a narrow separation induced by, say geography, could easily result in allopatric speciation along contemporary continental bounds, kind of like the famed Grand Canyon squirrel speciation example that is often cited in high school biology books. Of course, it is important to note that the time intervals preserved on each continent or within a given depositional basin are rarely the same, so we don't always have a precise apples-to-apples comparison, but as of right now, metoposaurids are no longer viable for global biostratigraphy. New horizons Despite an impressive surge in metoposaurid work within the past decade (more than 20 papers focusing exclusively on metoposaurids, with many others including them in a broader sample), there remains a lot of work on all fronts for metoposaurids. We outline many of these in the paper, and I'll list a few of them below. Phylogenetic grass The phylogeny of metoposaurids remains utterly unresolved, which is not really that surprising when considering the limitations of morphology-based phylogenetic inference for any clade and the stark conservatism in metoposaurid morphology. Through a series of differnet analyses, we demonstrated that resolution is highly sensitive to minor differences in algorithms, analytical methods, and scoring philosophy such that it's quite hard to be confident in most nodes. Certainly I think that people could do more testing (there are a million different combinations of settings), after the first set of computer-assisted phylogenies from Chakravorti & Sengupta (2018); Buffa et al. (2019); and Gee et al. (2019), this one has pretty clearly demonstrated that we are going to need a lot more data (i.e. more fossils) to even have a chance of producing any topology with measurable support for recovered nodes. 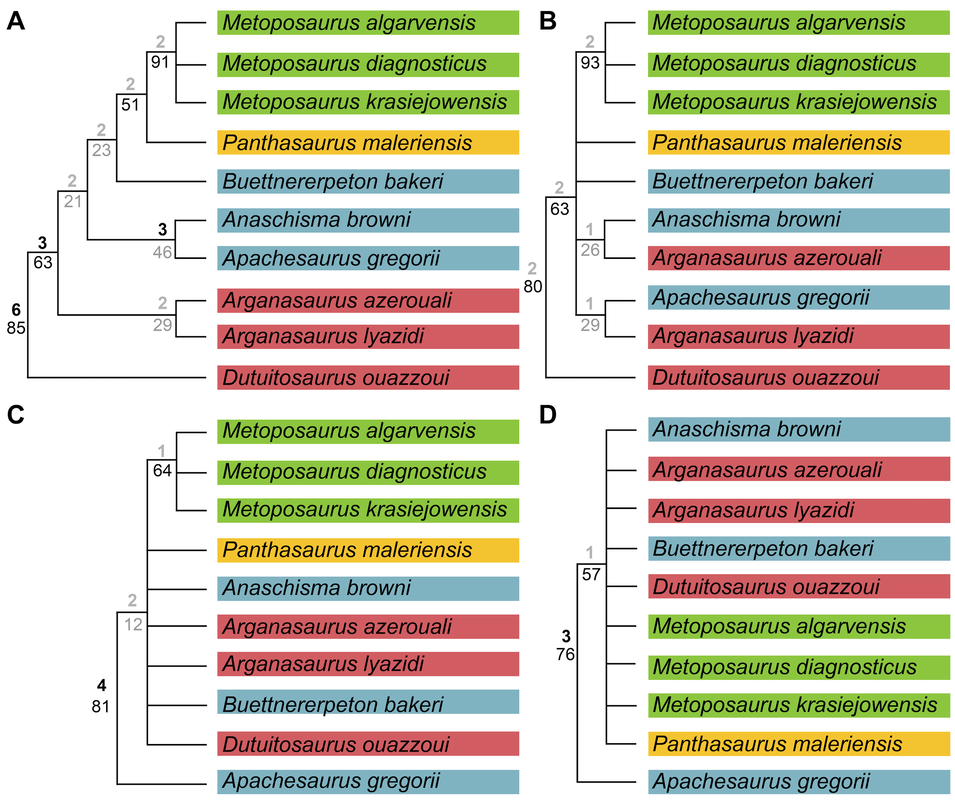 Comparison of phylogenetic hypotheses of Metoposauridae: (a) original topology recovered by Buffa et al. (2019); this was far more resolved than any other previous study; (b) reanalysis of Buffa et al.'s matrix with ordering of certain characters; (c) reanalysis of Buffa et al's matrix with some scoring changes but no character order; (d) reanalysis with both scoring changes and some character ordering. Apples and oranges One of the often unstated limitations to the metoposaurid record is that most taxa are only known from what are probably already adult individuals. We know nothing of whether there was a larval stage in metoposaurids or of what ontogeny looked like at skull lengths less than 10 cm (metoposaurids could reach at least 65 cm or so in skull length). One of the intriguing yet oft-overlooked aspects of this incomplete record, which we illustrate below, is that one of the lesser known metoposaurids, Arganasaurus lyazidi from Morocco, is only known from small specimens. This attribute is much better known and discussed for Apachesaurus gregorii, but although the describer of Ar. lyazidi, Jean-Michel Dutuit, speculated (in French) that it might be a dwarfed taxon, this has never been adopted by other workers, let alone tested. It's interesting to juxatpose this against the growing acceptance of my proposal that Apachesaurus is also not a dwarf taxon, although certain workers continue to advocate this while ignoring all of my previous studies (likely because they have no way to refute the histological data and related arguments). How the disparity in distribution of known specimens might in general affect our interpretations of metoposaurid taxonomy (are some purportedly diagnostic features really ontogenetic features) remains to be well-explored and arguably requires someone to get lucky and find some breeding pond or something of the sort. 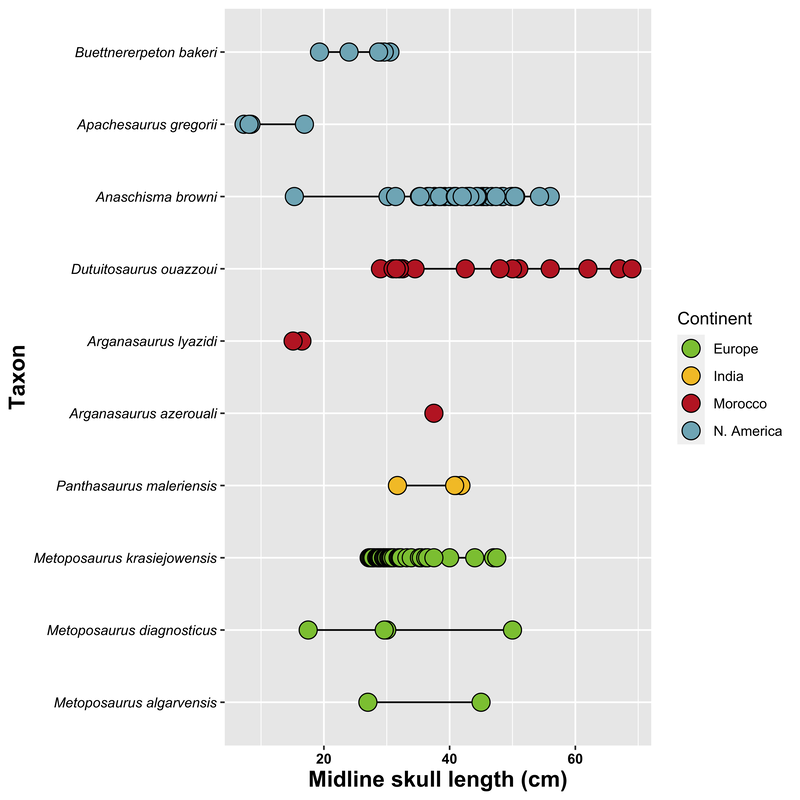 Old horizons One of the downsides of being an extremely common taxon is that people rarely collect your fossils. Metoposaurids are so abundant that there is an unspoken predisposition against spending a lot of time collecting metoposaurids, which are on the larger side of the Late Triassic tetrapod scale and definitely on the upper end of the most abundant group known from the Late Triassic of North America. One of the problems is that this leads to a lot of poorly documented reported occurrences, which get dodgier as you work down to a particular genus or species. Most reports of North American metoposaurids are asides in faunal lists or review papers, and they don't provide photographs, description, or even specimen numbers. As a result, a lot of these "reports" are not reproducible. Particularly in North America, this translates into irreproducible stratigraphic ranges - what are the highest and lowest occurrences of the given taxa. In some instances, there is good documentation of the fossils, and it's the sites that are just not well-constrained stratigraphically (this is the case for B. bakeri's three localities), but for Anaschisma browni and Apachesaurus gregorii, which have thousands of specimens referred to each of them, it's more often that the stratigraphy is reasonably well-resolved, and the taxonomy is suspect (or at least not demonstrated). The North American taxa are particularly susceptible to circular logic where both size and stratigraphic position are used as identifying features in the absence of any actually diagnostic anatomical features. 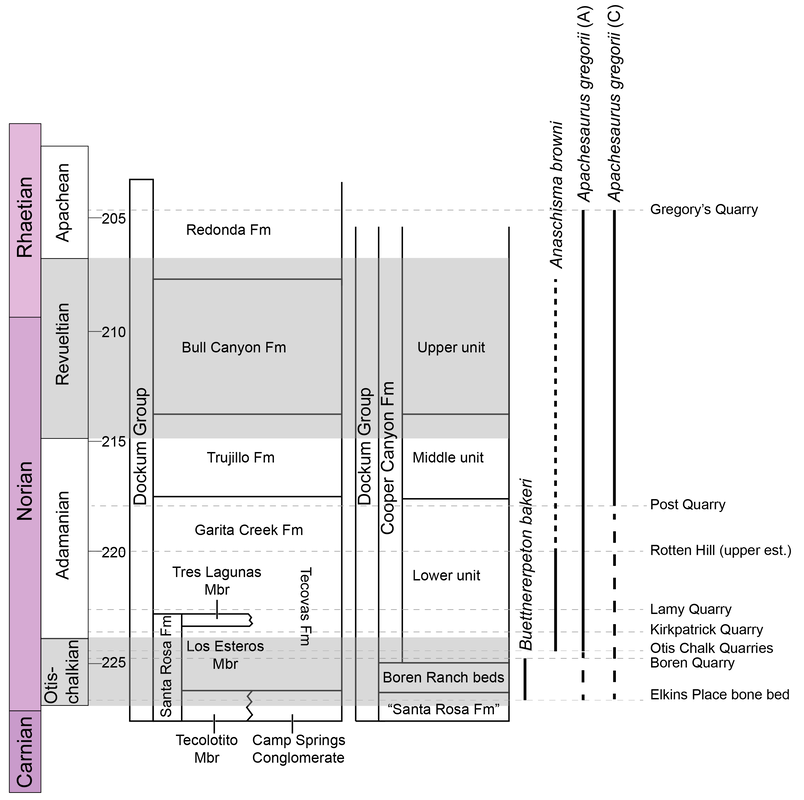 Stratigraphic range and uncertainty of North American metoposaurids. The two different Apachesaurus ranges represent all referred material (A) and only diagnostic cranial material (C); this relates mainly to whether small, elongate intercentra can be referred to Apachesaurus in isolation (I say no, so I favor range C). Dashed lines indicate uncertainty. The outcome of this is extreme uncertainty in what are the actual stratigraphic ranges of these two taxa, indicated above by dashed lines. The debate over Apachesaurus and whether small, elongate intercentra are really diagnostic for the taxon or just a hallmark of early ontogeny in North American metoposaurids has a huge impact on both the total abundance of specimens and the stratigraphic range of this taxon. Therefore, establishing the true range of these taxa will require a huge undertaking that involves both revisiting of specimens thought to be near the upper and lower bounds to determine whether they are diagnostic and revisiting of the sites that they come from to determine whether they are actually where previous workers have situated them. TLDR: there is still a lot of work to be done on North American metoposaurids, please collect them and then study them!
David Marjanović
10/12/2022 03:35:53 pm
Congratulations!!! Comments are closed.
|
About the blogA blog on all things temnospondyl written by someone who spends too much time thinking about them. Covers all aspects of temnospondyl paleobiology and ongoing research (not just mine). Categories
All
Archives
January 2024
|
 RSS Feed
RSS Feed
