|
To kick off this topical series looking at how we know what we claim to know about temnospondyls, we'll start with the most recognizable part of any animal: the head! Lots of important things happen up at the head - breathing, eating, and plenty of sensory functions from smell to hearing. So it stands to reason that the skull might offer us a good deal of insight into the lifestyle and ecology of temnospondyls. Shapes and sizes
The first is in relation to the snout: long- versus short-snouted forms. Of course, this begs the question of what really is a snout. Do all animals have snouts? We often associate snouts with mammals and many reptiles, but what about other animals? Do birds have snouts? Or just beaks? No modern amphibian has a long skull today, so do any of them have snouts? What about fish? Well, there is no standard, which is just like most other things in paleontology, but most people use 'snout' to refer to what is technically called the 'preorbital region,' or the amount of skull that is in front of the eyes. In this sense, all animals have a snout, it may just be really short in some.
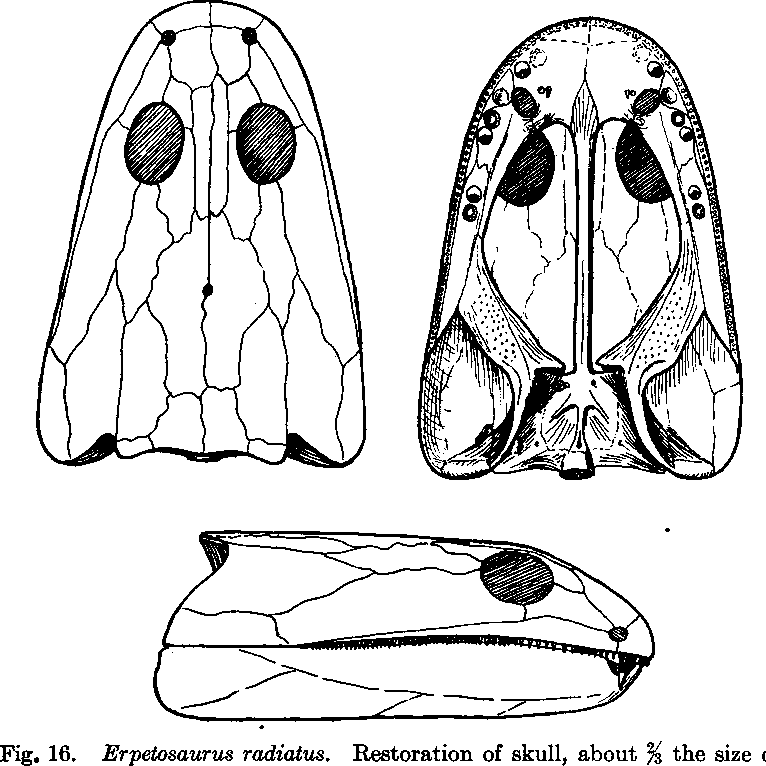
Short- and long-snouted should not be conflated with short-skulled (brachycephalic) and long-skulled (dolichocephalic); a taxon can have a long skull but a short snout, as with Erpetosaurus below (from Romer, 1930). Probably the closest thing to a taxon with the inverse - a short skull but a long snout - is a zatracheid, like Acanthostomatops below (from Witzmann & Schoch, 2006).
Most of what we infer about general skull shapes' relevance for inferring paleoecology is based on what we have today. So we assume that long-snouted taxa like Archegosaurus were crocodile analogues and probably spent more time in water, at least when it came to feeding. Other animals with long snouts that hang out in water are obligately aquatic (i.e. they die if they're out of water) like dolphins/porpoises and some fish like gar and needlefish. There aren't many terrestrial animals with long snouts, and those are weirdos with specialized lifestyles like anteaters and echidnas who use their snouts to get into places where termites and ants live. A cautionary tale comes in the most relevant case study: modern amphibians. There are no long-snouted modern amphibians, let they live in all sorts of different habitats. 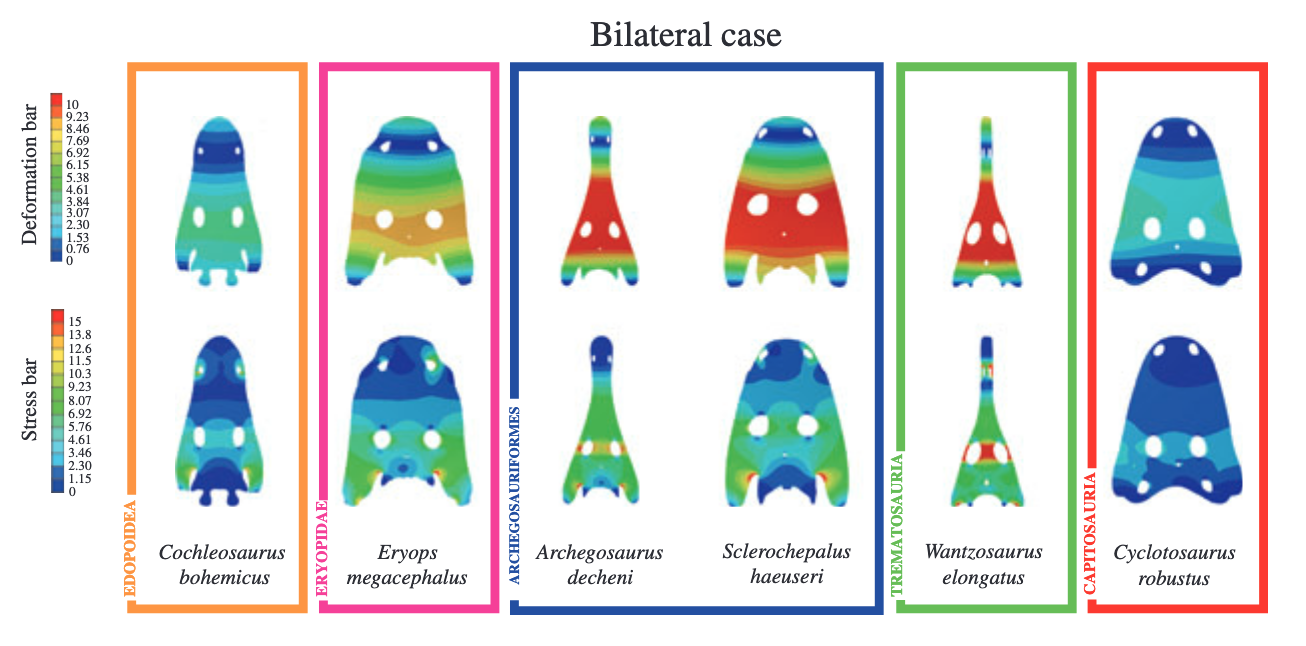 One promising type of analysis that has been applied to the entire skull to infer feeding behaviour is called finite element analysis (FEA). This method measures stress buildup in different regions based on the type of stress applied (e.g., from one side or evenly distributed). A number of studies have used this method to examine different feeding strategies in temnospondyls (e.g., Fortuny et al., 2011, 2017), with the idea that strategies that produce high degrees of stress are unlikely to have been used. You can see in the above example that the same stress produces different stress distribution patterns in different skulls (warm colours = more stress). However, much of the determination about possible feeding style and habitat is influenced by nuances of the skull like the exact shape of the tabular horns (the backward projections), not just its general shape. For example, not all of the taxa that lack the long crocodilian-like snout are inferred as terrestrial here.
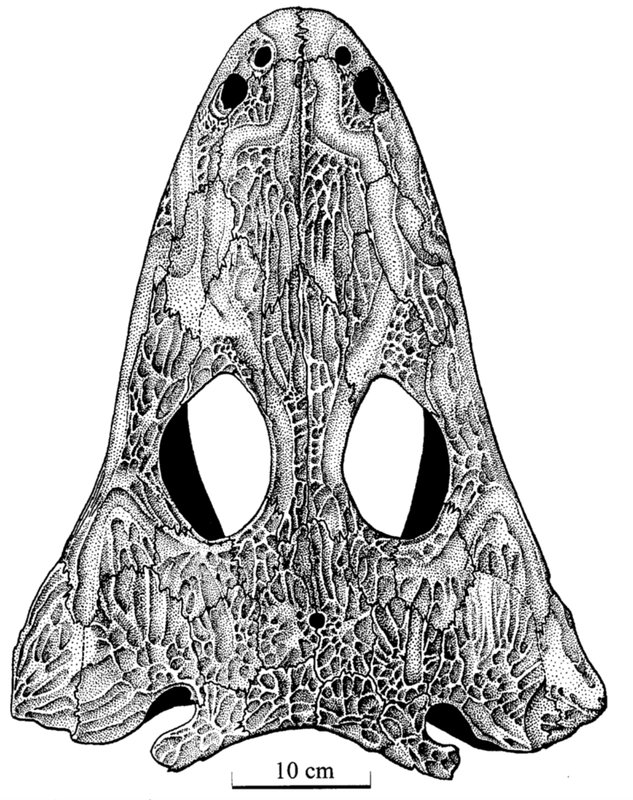
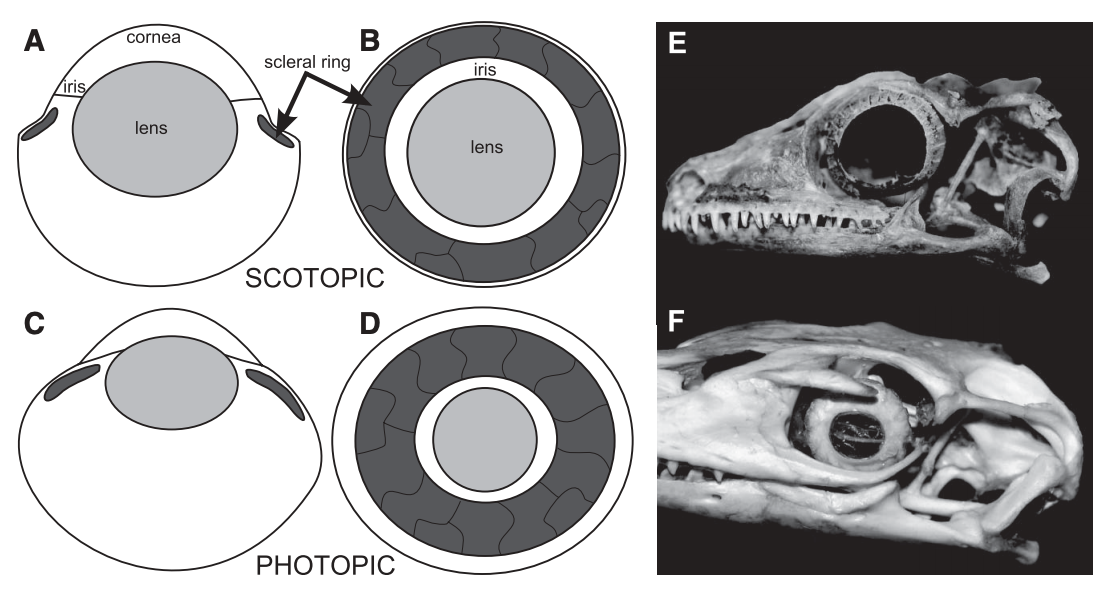
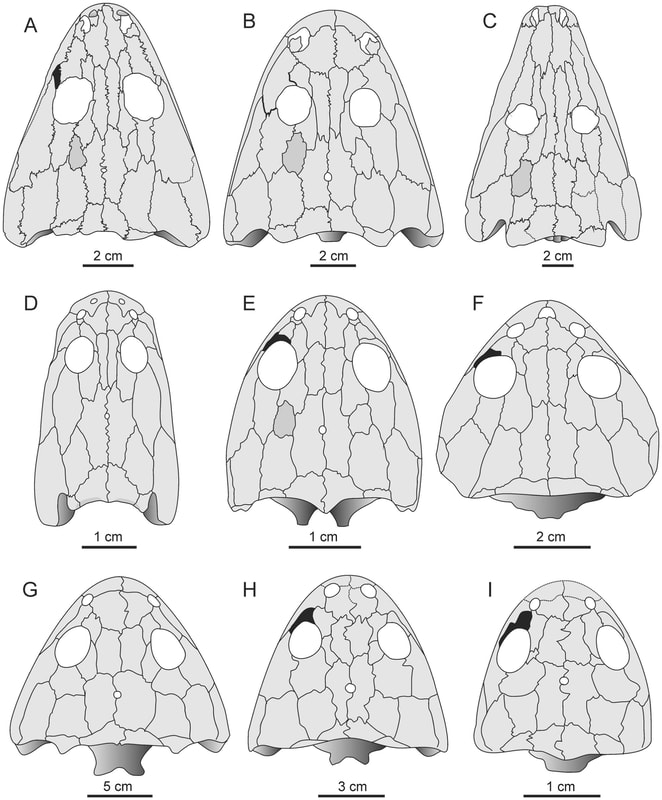
I spy, with my little eye...One of the general attributes of skulls that may confer pretty good insight is one of the primary sensory features - the eyes! Obviously, which way your eyes face is a major constraint on how well you can catch other animals, avoid predators, etc. Temnospondyls exhibit lots of variation in the position of the orbits (the eye socket), from facing mostly outward to the site to facing entirely upward.
In tune
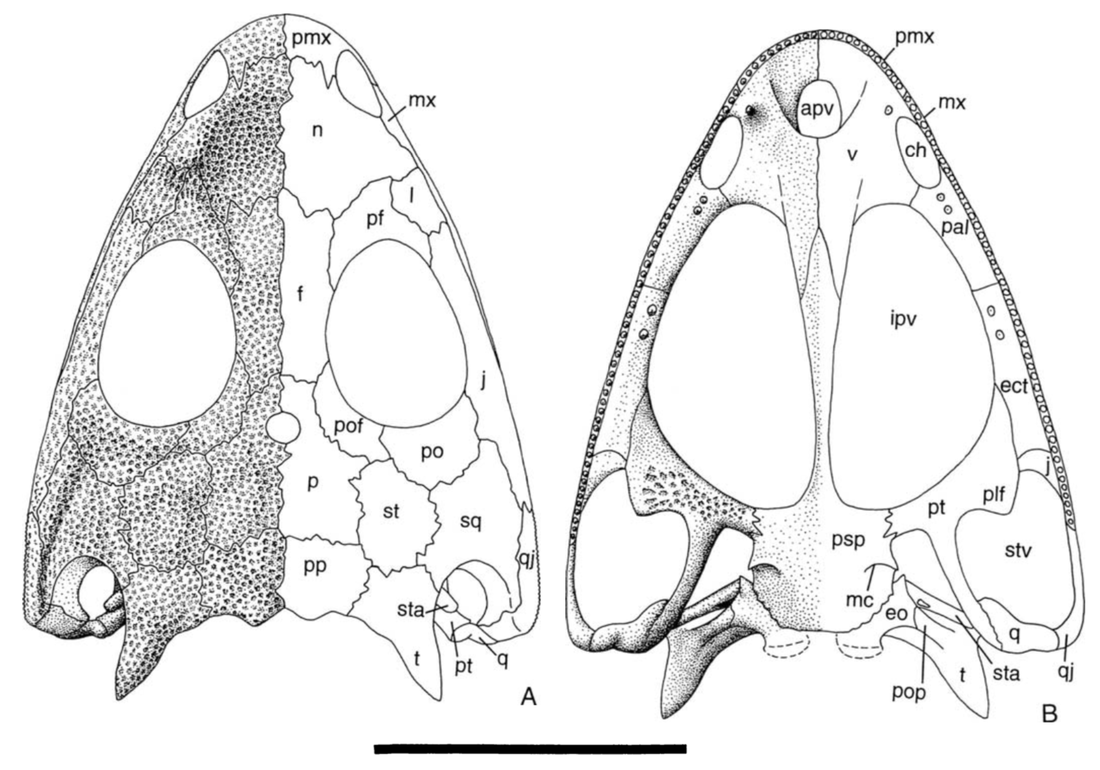
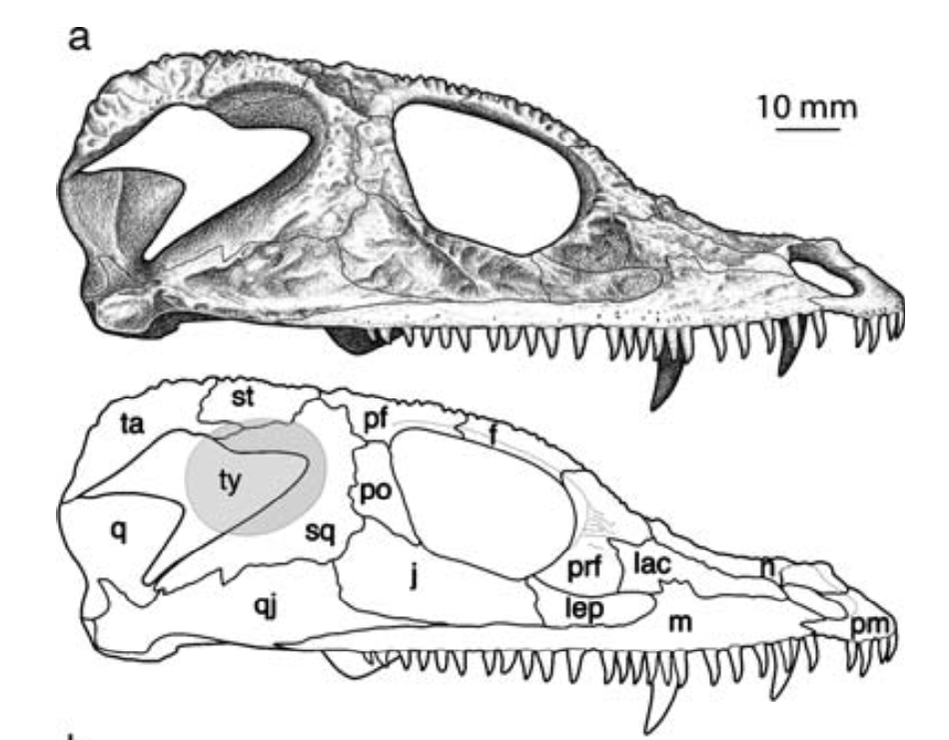
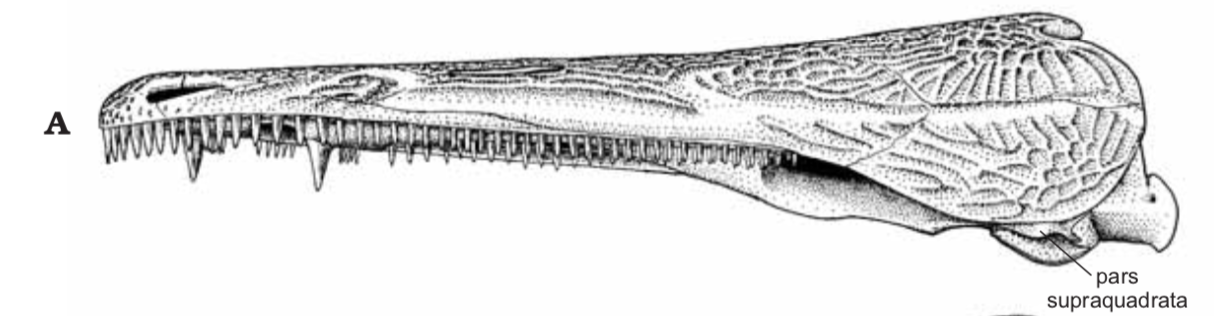
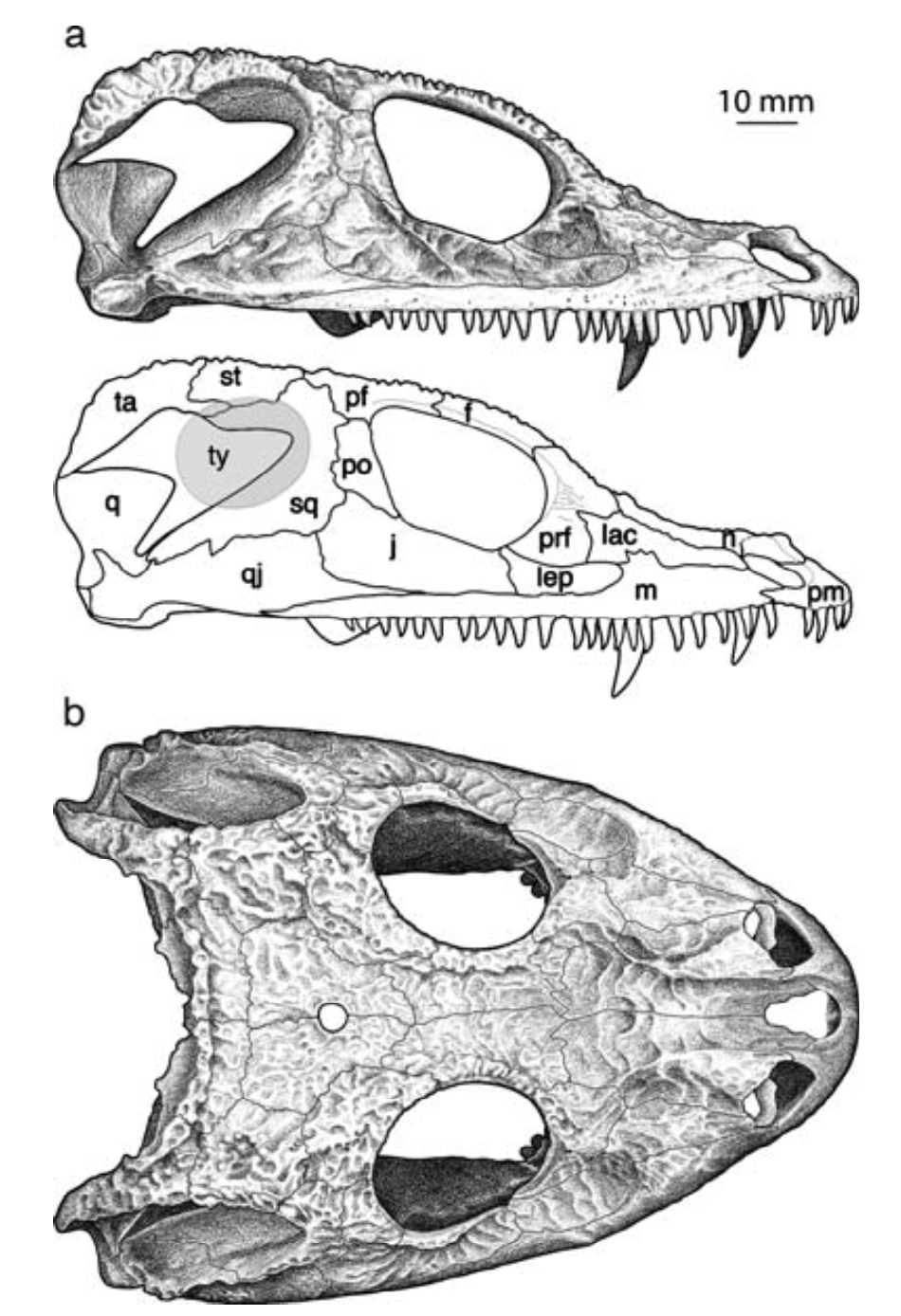
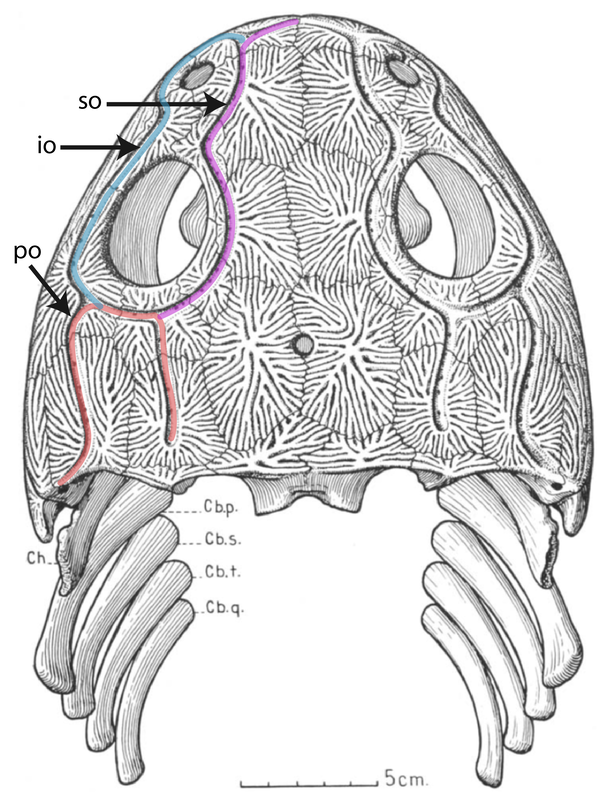
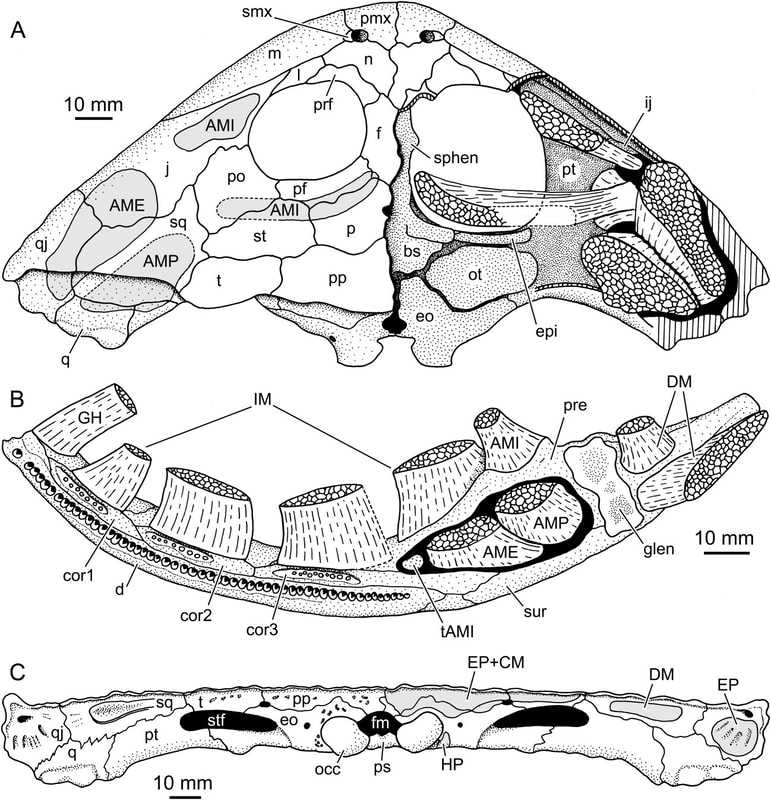
All of this is to say that it is quite hard to actually figure out what kind of auditory capabilities temnospondyls had and in turn whether a given species was better at hearing on land or underwater. We know that at least some temnospondyls had a tympanic membrane, which suggests that they were capable of detecting airborne sounds and thus probably spent at least some time on land. This evidence comes from a well-developed otic notch, a large opening in the skull behind the orbit and adjacent to the braincase. Whereas most of the temnospondyl skull is covered in pits, ridges, and grooves, this otic notch sometimes has a large unornamented flange in it that is thought to have been the spot where a tympanum would have attached (the gray shading and 'ty' label in the figure on the bottom left). Often times, one of the bones of the inner ear (the stapes), is preserved with one end pointing outwards into the notch and the other end articulated with the braincase. This suggests that these temnospondyls could hear airborne sound.
However, most temnospondyls have an otic notch, and the majority of those do not have this large flange within their otic notch - many have no flange at all, particular those in which the otic notch doesn't face out to the side, but upward. Species without a flange do still have a stapes that points towards the notch, but the key is that it is unknown whether a tympanic membrane stretched across the otic notch or whether it was merely covered in other soft tissues like the rest of the head. It has been suggested that the otic notch in these taxa was indeed covered by some soft tissue and thus formed an enclosed air pocket that was impermeable to water. If the stapes projected into this pocket, then it would be operating in an airborne setting, not an underwater setting, so vibrations transmitted through water would be converted to sound waves inside of this air pocket and then be conducted to the brain in the same fashion as animals with a tympanum. This is the best guess for what many of these taxa with otic notches but no tympanum were doing (e.g., Schoch, 2000; Arbez et al., 2017).
Slimy senses
Something to chew on The jaw sadly does not tell us much about temnospondyl ecology so much as what sits on the jaw does: the teeth. Temnospondyls do have a variety of different jaw anatomies, and it's clear that some features, like the size of the cavity where muscles would have been packed in or the length of a long protrusion off the back end (the retroarticular process), have functional implications related to bite force, gape (how big the mouth can open), and jaw-opening mechanics (did the head raise up or did the jaw lower). Most of the studies that look at the role of the jaw do so only in a conjectural sense of attempting to reconstruct the arrangement of muscles (e.g., Witzmann & Schoch, 2013) or a modeling sense of attempting to predict where stress would have built up during a bite (e.g., Fortuny et al., 2011). Early histological work has identified complex articulations between different bones in the jaw (e.g., Gruntmejer et al., 2019), some of which are set up for tension (pulling apart) and others of which are set up for compression (pushing together), but we don't have enough data yet for comparative purposes to really know what this means other than that some parts of the jaw were under different stress than others (this is true of pretty much any animal's jaw). There is clearly a lot of room to work on jaw biomechanics because at present, we pretty much assume a lot of things didn't vary much between temnospondyls.
Tale of the teeth One would assume that the teeth of temnospondyls should tell us quite a lot about what they ate; after all, this is often how we infer what modern animals are up to. Alas, the teeth of temnospondyls are about as basic as they come. Practically all temnospondyls have the stock model of tooth: a single cone. This type of unspecialized dentition is common of many fish and early tetrapods, although there are always exceptions that end up being unusually specialized. Many modern amphibians also have only conical teeth. One thing that we know that conical teeth are good for is eating fish; cones are great for piercing and stabbing prey, and something slippery like a fish that's hard to hold often requires a firm grasp. You can see this in many modern predatory fish as well as in crocodilians (which of course eat pretty much anything, not just fish).
This is not to say that every temnospondyls with conical teeth (the overwhelming majority) were fish-eaters. Some of them were almost certainly too small to be eating fish unless they were eating guppies, and because amphibians lack true necks, it would be difficult for many land-dwelling temnospondyls to eat much of anything in the water. There are also nuances to conical teeth - some area straight while others are curved; some have cutting edges while others are entirely smooth. One might assume, for example, that the ones with cutting edges are eating some beefier items (say, other temnospondyls). Most are not perfect cones. Size differences were probably also a big factor, mainly in controlling the number of teeth that fit in the mouth - some taxa had fewer than 30 teeth along the outer margin, while others exceeded 130.
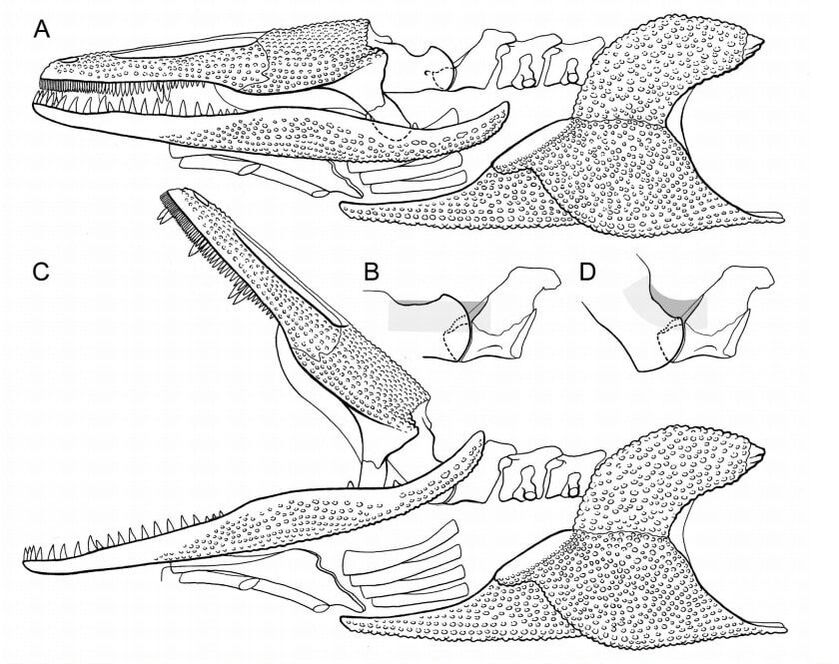
Finally, we have teeth in different parts of the mouth, which is a strange concept for people to think about (you can read more about the very large teeth of many temnospondyls here). In addition to the outer tooth row ('marginal teeth'), temnospondyls almost always had large accessory teeth on the roof of their mouth that we call 'fangs' or 'tusks.' Whether these were used to impale prey isn't clear; the bone under these teeth is often quite thin, so it may have been pretty bad at bearing a lot of bite force on those teeth. Then we have very small teeth, often found by the hundreds, called denticles. These covered the roof of the mouth in early temnospondyls (like Cochleosaurus, above on left; Sequeira, 2003) but gradually become reduced and eventually disappear in later temnospondyls (like in Mastodonsaurus, above on right; Schoch, 1999). Presumably, having the roof of your mouth was useful for prey capture by making a giant gripping surface (imagine a sandpaper-like covering). This would be great for slippery prey or anything that would try to make an escape from the literal jaws of death. So it isn't clear then whether the loss of these denticles had any measurable effect on a temnospondyl's ability to eat, and if so, why these tiny teeth were lost. There is no clear correlation with any other skull feature (e.g., being short-snouted) that might indicate a particular need for denticles in one type of environment versus another. That wraps up what was a much longer blog post than I anticipated (and which is hardly exhaustive). The TLDR is that there are few simple ways to infer a temnospondyl's ecology based on singular features, like whether it had a wide head or curved teeth. We can make educated guesses of course (and often do), but most of this is based on the limited sample of what we can parallel to today, like crocodiles or frogs. We could be entirely wrong for all we know! That's why it's important to look at all parts of the skeleton, not just the head. Up next week: limbs!
David Marjanović
10/16/2021 07:24:16 am
Yay! Another post! And such a long one! ^_^ Comments are closed.
|
About the blogA blog on all things temnospondyl written by someone who spends too much time thinking about them. Covers all aspects of temnospondyl paleobiology and ongoing research (not just mine). Categories
All
Archives
January 2024
|











 RSS Feed
RSS Feed
