|
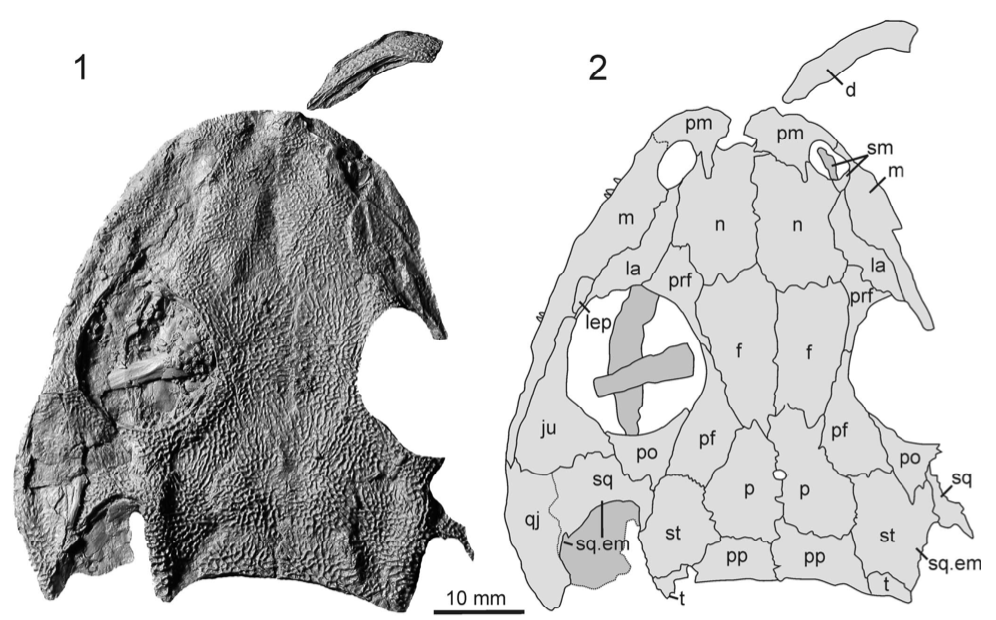
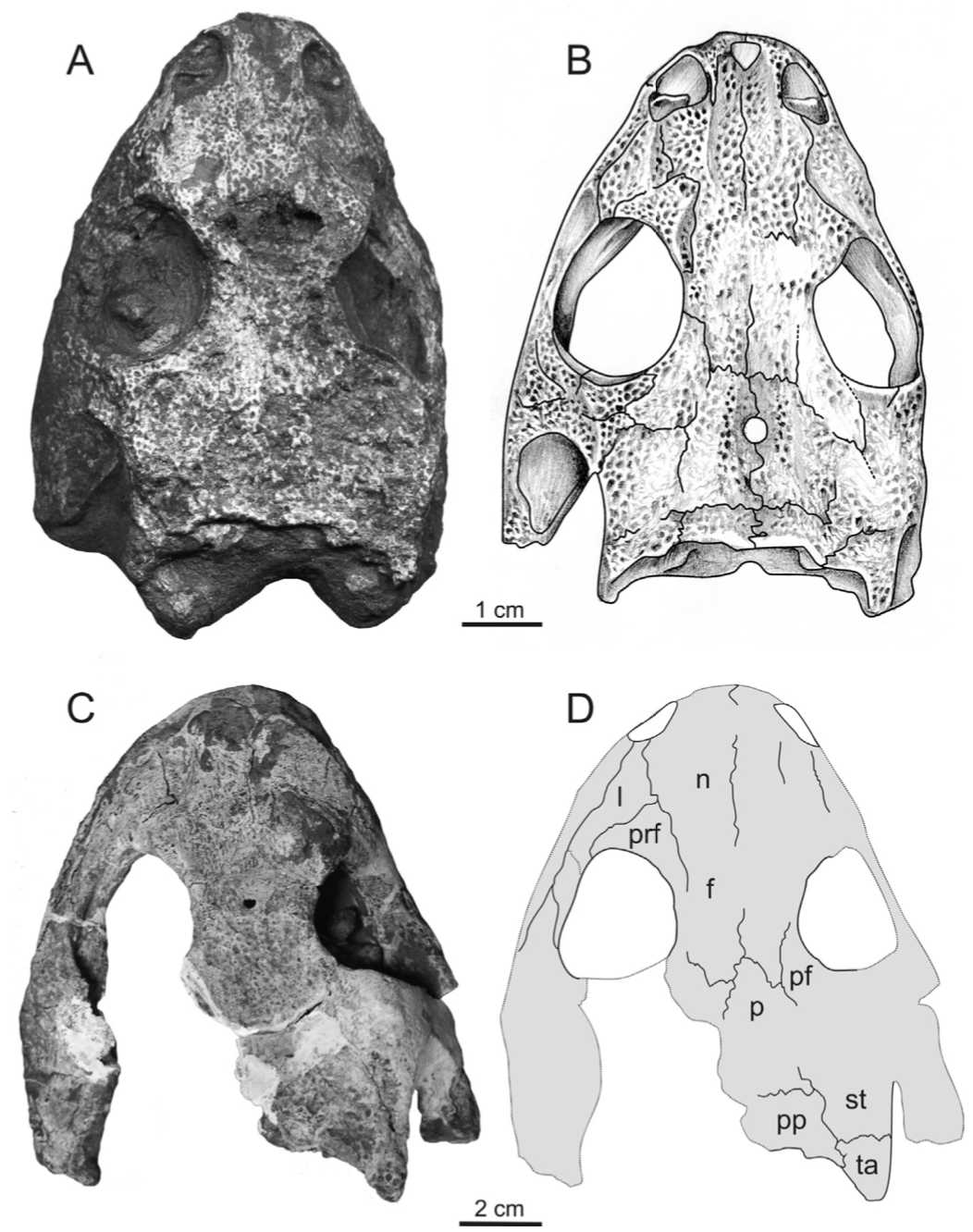
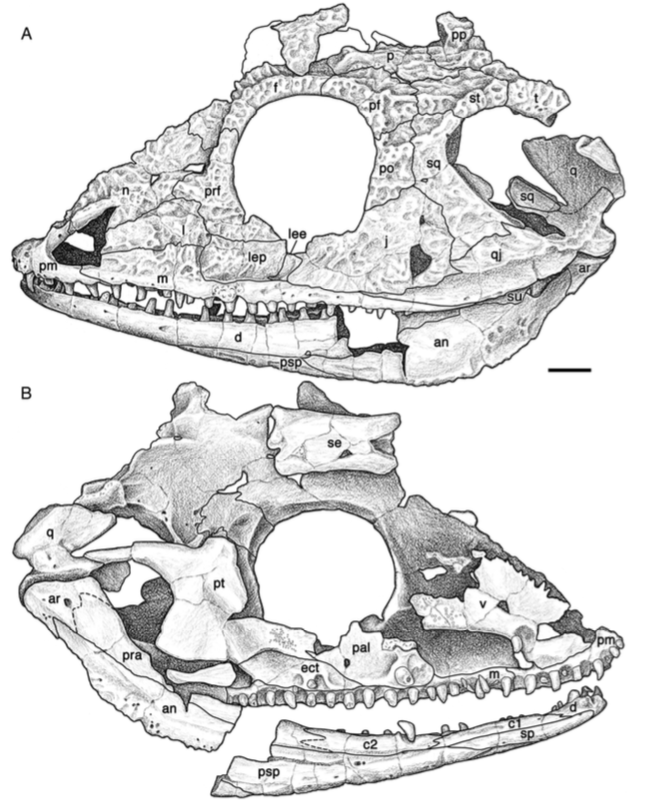
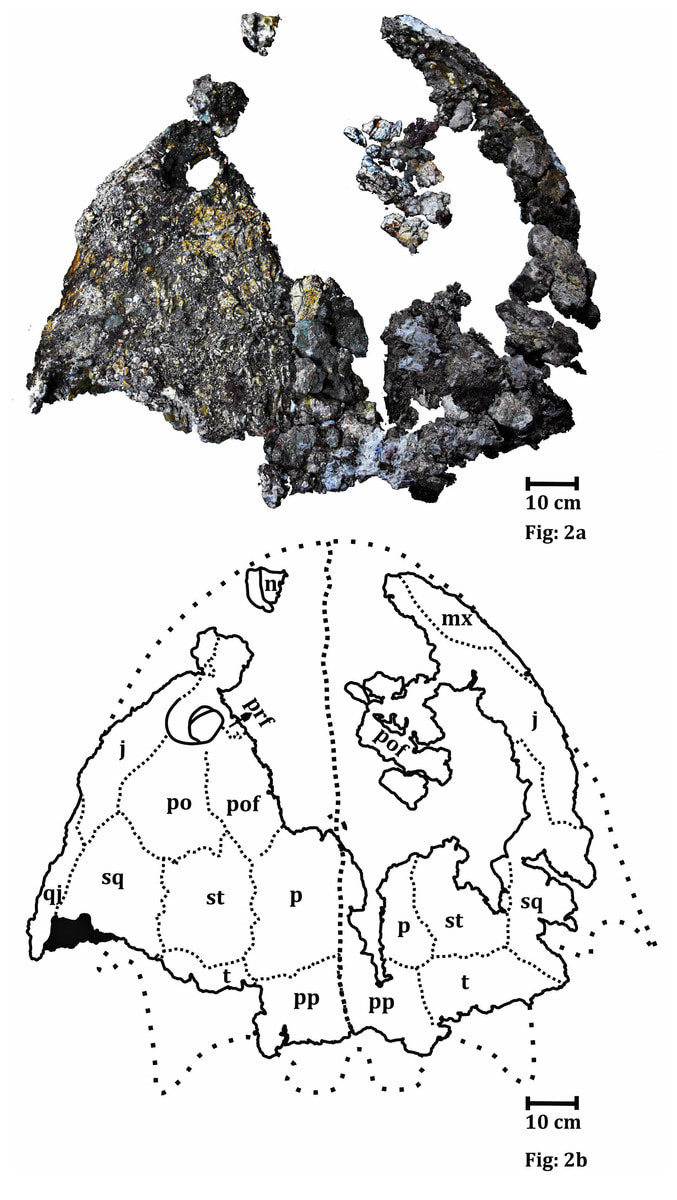
Title: Description of the metoposaurid Anaschisma browni from the New Oxford Formation of Pennsylvania Authors: B.M. Gee, S.E. Jasinski Journal: Journal of Paleontology DOI: 10.1017/jpa.2021.30  The amazing art above was done by Sergey Krasovskiy; you can find his DeviantArt profile here and his Twitter here! The number of metoposaurids is not random - we know that there were at least two individuals preserved at the site based on the number of duplicate elements from the site. General summary: This study provides the first detailed description of metoposaurid temnospondyls from Late Triassic deposits in Pennsylvania. Metoposaurids are well-known for their widespread distribution in the Late Triassic, including in the southwestern U.S., but they have a much scarcer record west of Texas. The east coast in particular was geographically closer to other regions that preserve metoposaurids that have drifted apart over the millenia, like northern Africa and western Europe. The east coast is also the only region in North America where a large-bodied temnospondyl that isn't a metoposaurid is known from (this pattern of metoposaurid exclusivity is not observed anywhere else with decent sampling). While previous people have mentioned the Pennsylvania material and ID'd it, they never provided good figures or explicit justification for their ID, so essentially any reader was left to take them at their word. We formally describe the material, identifying it to Anaschisma browni (best known from Arizona, New Mexico, Texas, and Wyoming), which is not the same species that a metoposaurid from Nova Scotia belongs to, and discuss the importance of providing proper documentation and justification for taxonomic identifications (which are fluid hypotheses, not static facts). While the east coast is much more fossil-poor (this is definitely the best metoposaurid material from anywhere in North America west of Texas), it may also be the only region that preserves the transition from a diverse large-bodied temnospondyl assemblage to one that is entirely dominated by metoposaurids.
Title: New information on the dissorophid Conjunctio (Temnospondyli) based on a specimen from the Cutler Formation of Colorado, U.S.A. Authors: B.M. Gee, D. S Berman, A. C. Henrici, J.D. Pardo, A. K. Huttenlocker Journal: Journal of Vertebrate Paleontology DOI: 10.1080/02724634.2020.1877152
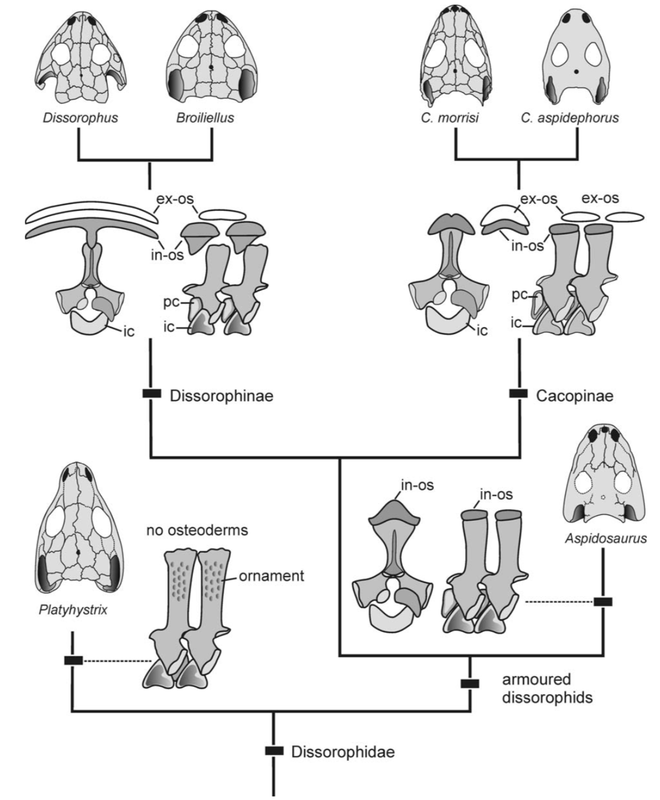
The Red RiverThe Red River is a mostly east-west oriented, eastward-flowing indirect tributary of the Mississippi River, stretching from the Texas Panhandle all the way to Louisiana (over 1,350 miles). It forms the border between Texas and Oklahoma (hence where the "Red River Rivalry" in collegiate football between University of Texas and the University of Oklahoma derives from), two of the most historically productive states for early Permian tetrapod fossils (especially the famed Texas redbeds). Much of our record of North American temnospondyls from this time comes from these two states, with more minor contributions from New Mexico, Utah, and Ohio. Dissorophids, the armoured dissorophoids, are among the best known taxa from these redbed deposits, and in many instances, the entire record of a given species is restricted to one or two states. Whether this is an artifact of the fossil record or an accurate representation of a truly restricted geographic range is often unclear; many taxa are in fact known from only one specimen or one site (the two species of Cacops below are examples; figures from Gee & Reisz, 2018 and Anderson et al., 2020).
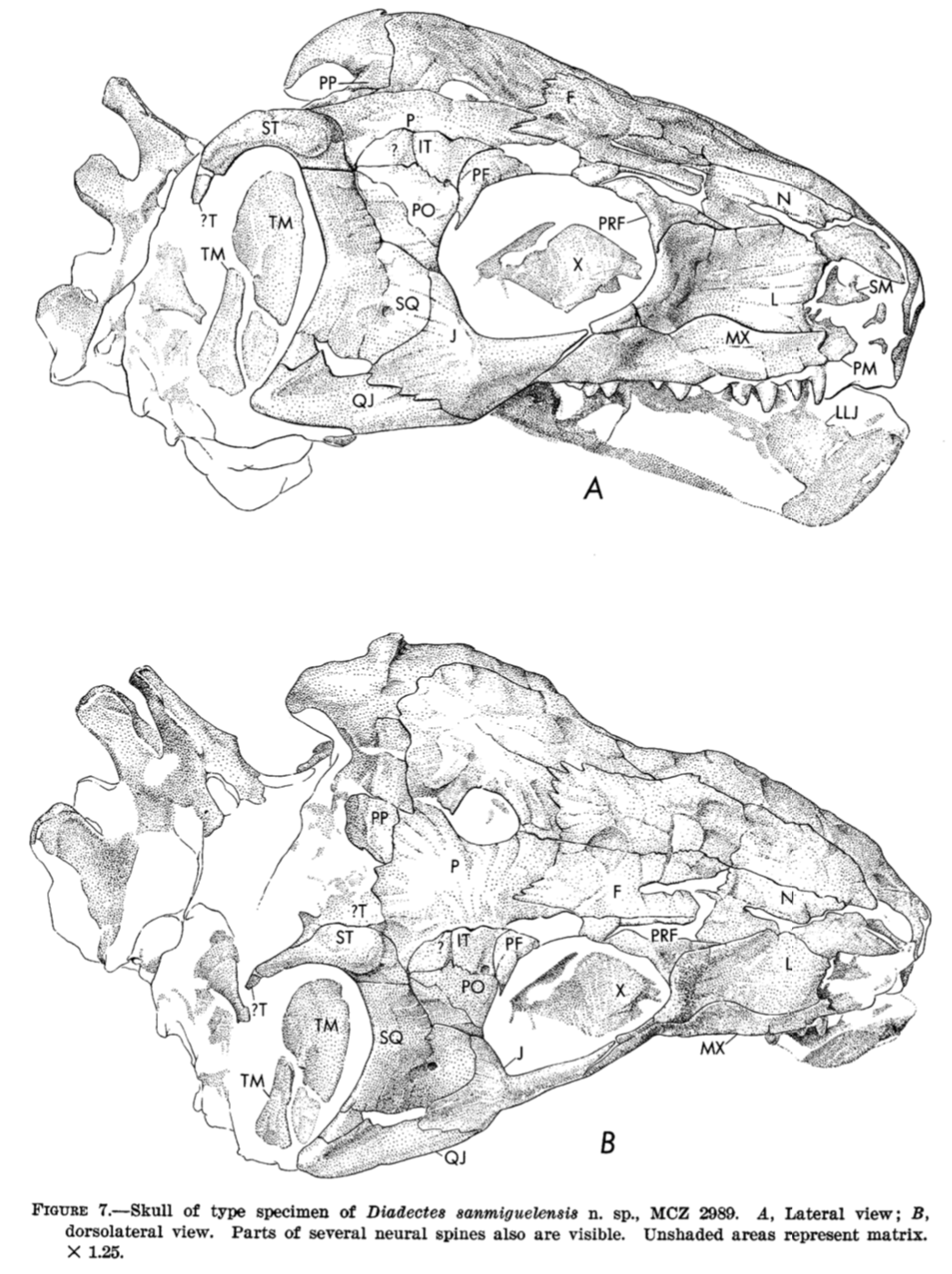
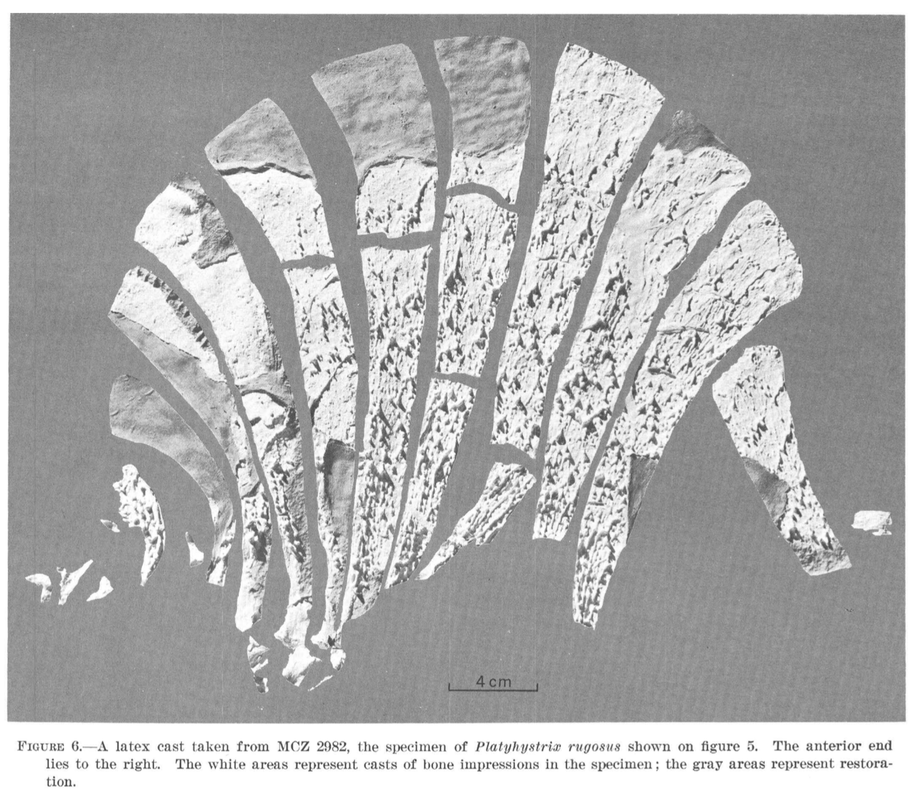
Land of Enchantment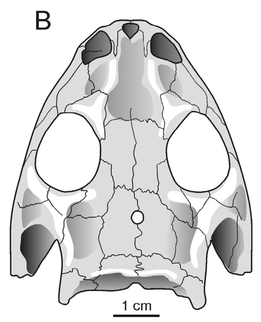 Conjunctio multidens (reconstruction from Schoch & Sues, 2013) is one such dissorophid, known only from two sites (but the same county) in New Mexico. Although New Mexico and Texas are adjacent states, their faunal assemblages were somewhat different, a result of regional geography no longer present that would have created some degree of spatial segregation. Other examples can be found in the trematopids Anconastes and Ecolsonia, only found in New Mexico, or the dissorophid Broiliellus reiszi, which differs starkly from other members of the genus that are known from Texas. Because the Texas redbeds have been so extensively sampled, with at least a dozen other dissorophids known, it can be reasoned that indeed Conjunctio also did not occur in Texas. But does that mean that it was only found in New Mexico? Not necessarily. The rest of the Four Corners region and the midcontinent is less well-explored, probably because the early Permian deposits that yield tetrapod fossils are less extensive. One of the well-known productive localities in the Four Corners region is in the Placerville area in Colorado, where the Cutler Formation (also found in New Mexico) is several hundred meters thick. First reported by Lewis & Vaughn (1965), the assemblage preserves one of the few sails of the dissorophid Platyhystrix (below on right), the holotype of Diadectes sanmiguelensis, a diadectomorph stem amniote (below on left), and a number of other tetrapods also known from other redbeds deposits. Synapsid fans may also know this as the type locality for the sphenacodontid Cutleria.
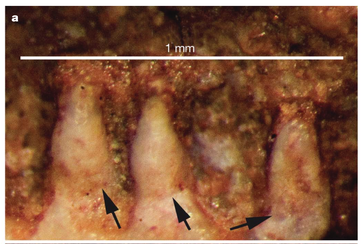
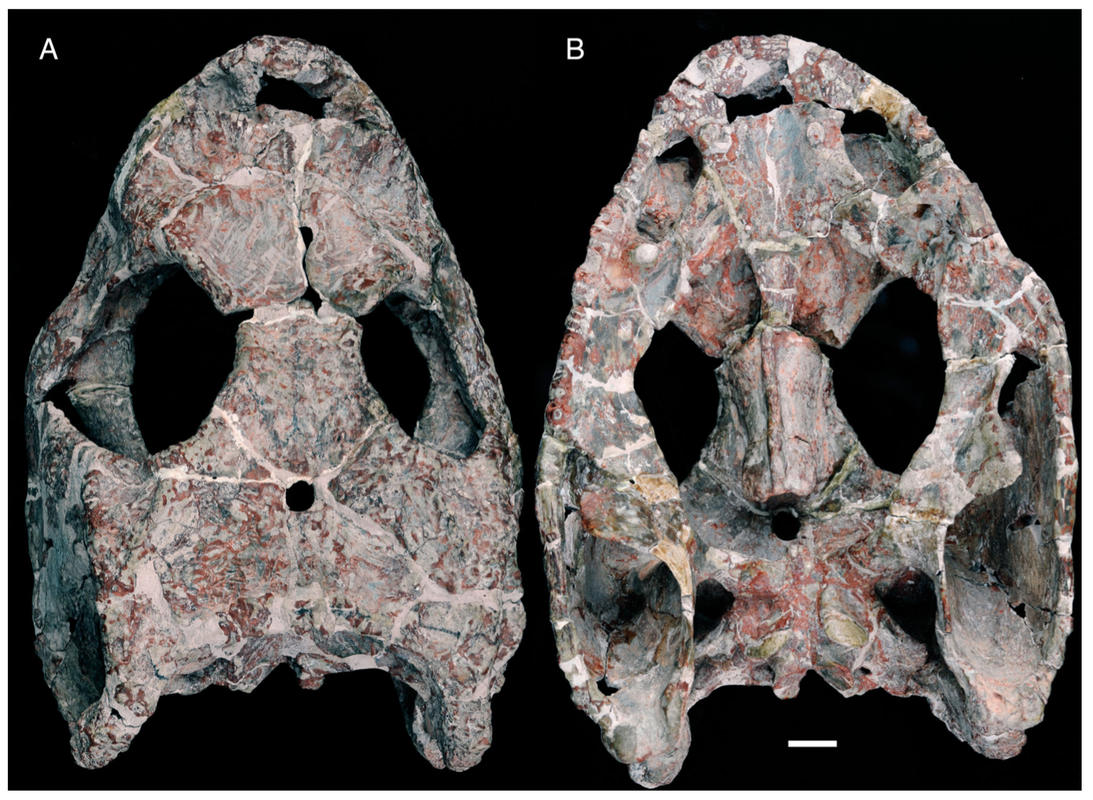
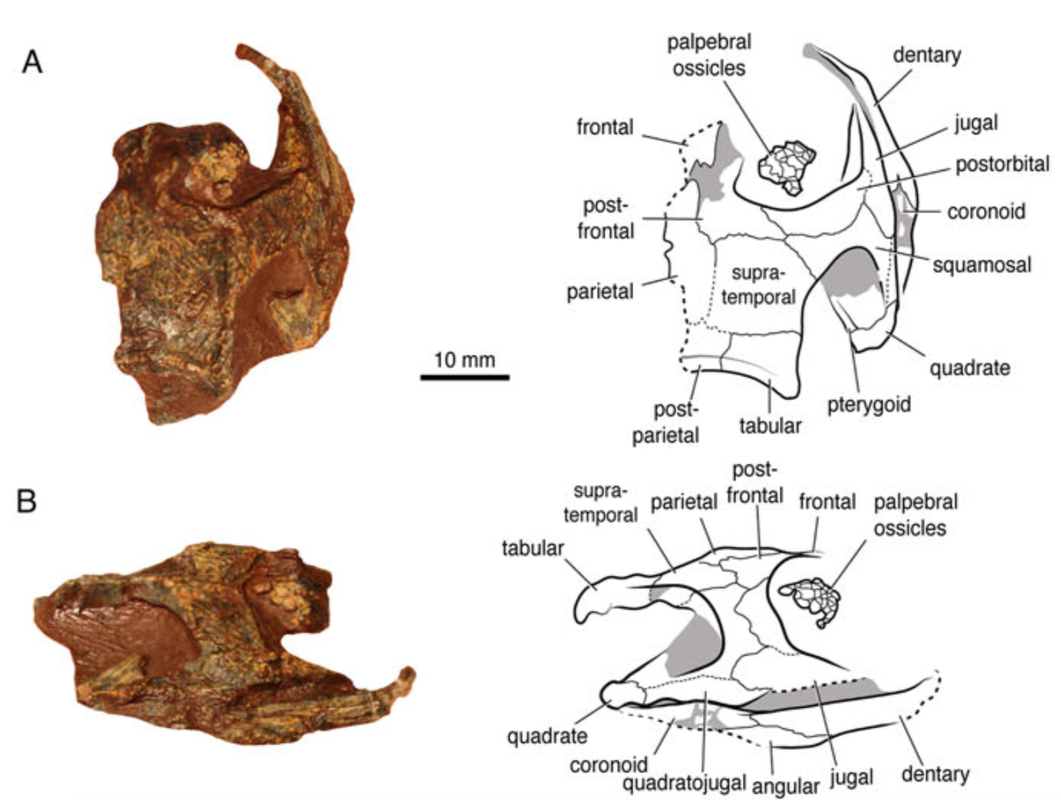
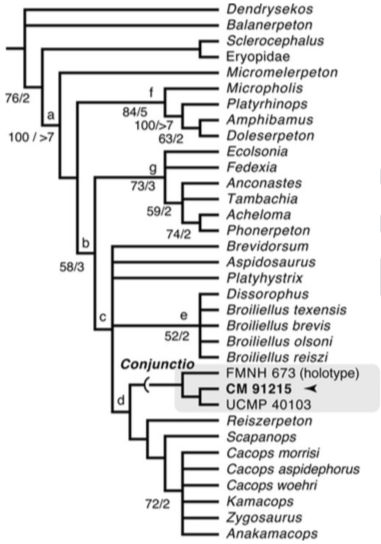
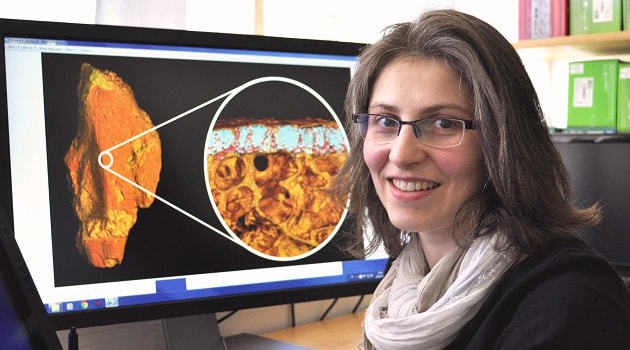
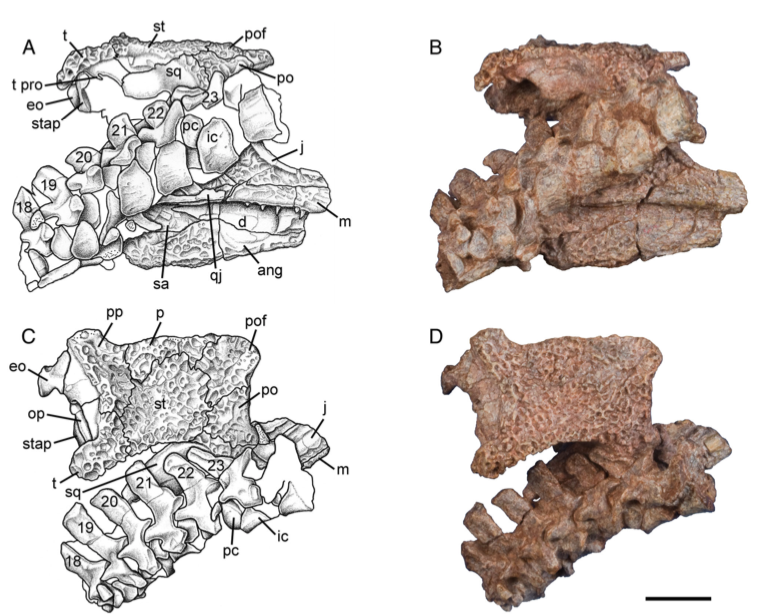
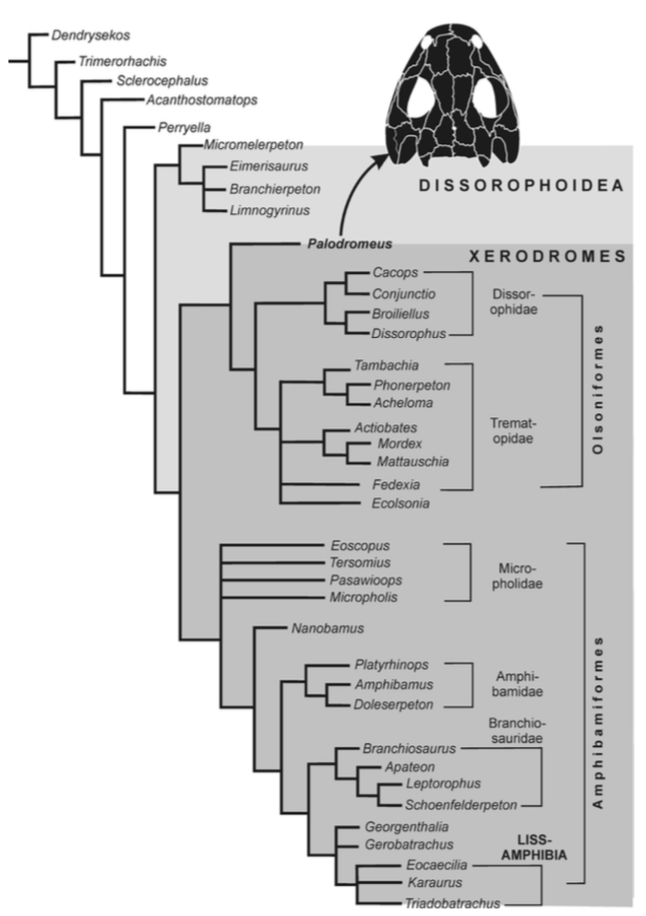
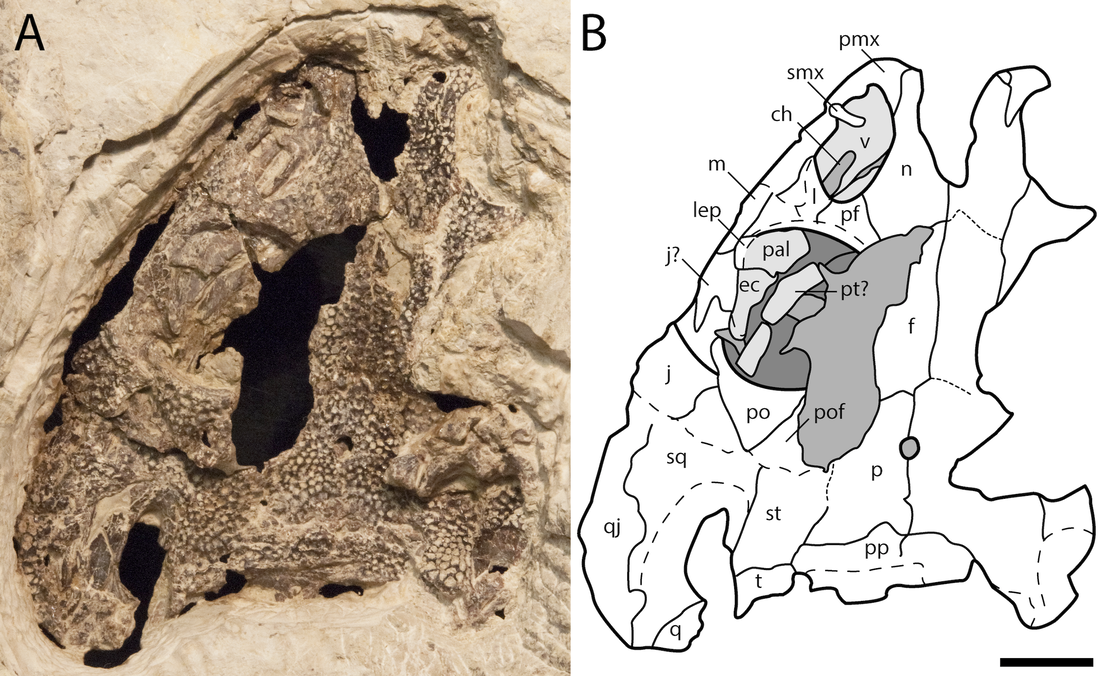
Terminology: While 'transitional form' or 'transitional fossil' is a commonly used term that even the general public knows (e.g., Archaeopteryx), scientists are moving away from it because it's misleading, suggesting that one animal directly turns into another (one of the most common misconceptions about evolution). In fact, this very rarely occurs (a process called anagenesis), and it does not capture major transitions like the theropod-bird transition or the fish-tetrapod transition. Instead, continual splitting of new species that go extinct is what gradually leads to a shift in a group's anatomy. A given species' suite of features may be transitional insofar as it captures part of this shift, but the species or individual itself is not. Agree to disagree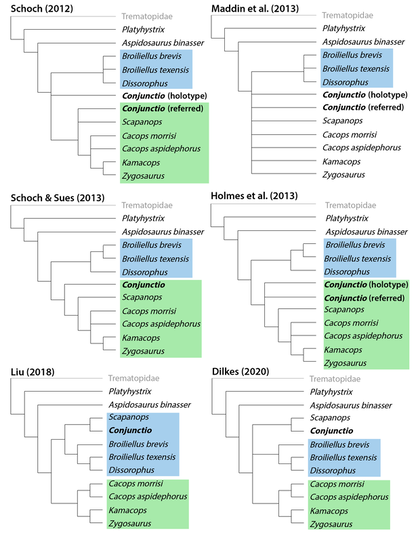 Comparison of different phylogenetic results with Conjunctio. Boxes refer to dissorophid subfamilies: green = cacopines, blue = dissorophines. Comparison of different phylogenetic results with Conjunctio. Boxes refer to dissorophid subfamilies: green = cacopines, blue = dissorophines. As one might expect from a taxon with a weird mixture of features, there is no agreement on the position of Conjunctio in phylogenetic analyses of dissorophids, as summarized on the right. Historically, the phylogeny did not support that they were conspecific (expected recovery as exclusive sister taxa), as they were frequently scored separately due to uncertainty over Carroll's referral (the referred specimen was called the "Rio Arriba Taxon"). It doesn't really help that the holotype (C-D) and the much smaller referred specimen (A-B) differ from each other in cranial proportions, somewhat questioning whether they are actually the same taxon (since Bob Carroll created Conjunctio in 1964). You don't have to be a paleontologist to look at the two specimens above and kinda wonder whether they are actually the same (or whether the bottom one can be said to be much of anything really). Sometimes both specimens were recovered as eucacopines (Holmes et al., 2013), sometimes only one was recovered as a eucacopine (Schoch, 2012), sometimes Eucacopinae didn't exist (Maddin et al., 2013), sometimes the composite was recovered as a eucacopine (Schoch & Sues, 2013), and sometimes the composite was recovered as a dissorophine (Liu, 2018). So basically every possible result short of it being recovered as a non-dissorophid! The Centennial StateThe new specimen that we report here was collected from what's known as the Placerville locality by my coauthors, Adam Huttenlocker and Jason Pardo, a few years back. It is not nearly as complete as the other two specimens, but it is well-preserved, enough to show some distinctive features not found in most or all of the other dissorophids. For example, the two bones called the postorbital (right behind the eye) and the supratemporal (a little farther back) usually touch, but here they do not (this is found in two of the species of Cacops as well). The jaw articulation (marked by the quadrate) also sits in front of the level of the back of the skull (marked by the postparietal); this is something found only in Dissorophus and Scapanops. So there is actually quite a lot of information in this little skull despite it being at best 25% complete!
Our phylogenetic analysis, which combines two previous matrices (it was originally intended for a larger sample), finally recovers not just the original two specimens, but also this third one as a proper clade (i.e. what you expect if they really do all belong to the same species)! The statistical support is not very good though, and it's quite possible that a future study could find a different result. Dissorophid phylogenetics remains very much in flux outside of just Conjunctio (something that I'm working on right now). COVID addendum: Lest someone think that all of us are superhumans who pumped out this paper in the pandemic, it was mostly completed before the pandemic really set in (we submitted the first version in May 2020). Jason and I in particular as current and recently graduated PhD students have really slowed down in putting new work into the pipeline during the pandemic, which I think is important to note since a lot of people are really struggling and should not be concerned with their perceived relative productivity! Refs
Inner workings: some fun (and not so fun) insights into demographics of temnospondyl research2/9/2021
Temnospondyls are a weird bunch of animals, which naturally means that the people who work on them are also a weird bunch. I've been kicking around some ideas for a blog post looking at the demographics of temnospondyl research for a while, both out of a general interest and with an eye towards diversity (or the lack thereof) in early tetrapod research, which is particularly niche among vertebrate paleontology. Meant to do it in 2020, but a lot of things obviously happened that delayed it to now... Where to find the temnos (Part 1)Where do temnospondyls "live" today? It's probably not that surprising that my semi-arbitrary list of the institutions with the most temnospondyl material are in predominantly western, northern hemisphere countries. 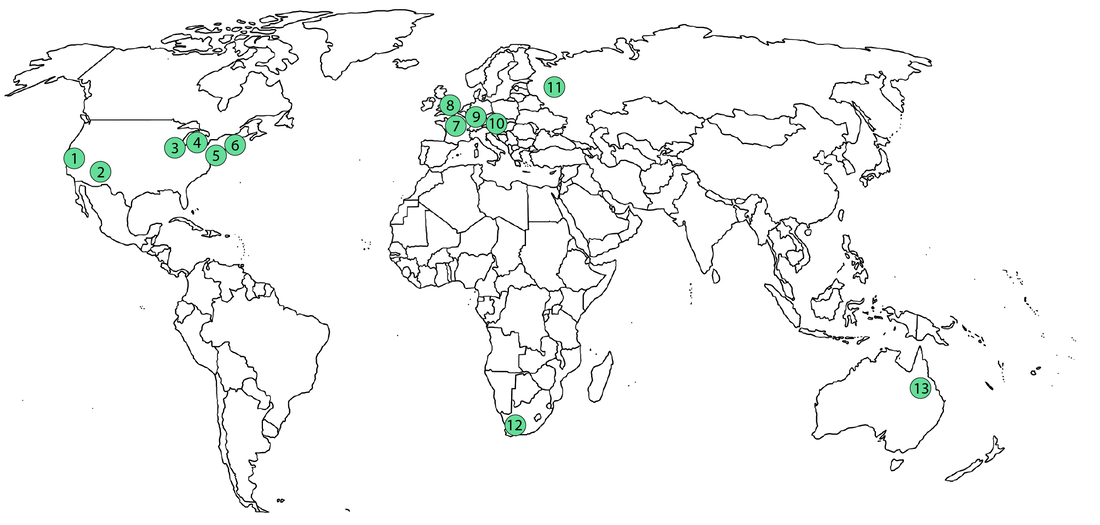 This is obviously not a full list; there are many other institutions in the U.S. with decent temnospondyl holdings (KUVP - Lawrence, KS; OMNH - Norman, OK), especially ones with large quantities of material from single localities (TTU - Lubbock, TX; PPHM, Canyon, TX), as well as a few international museums like BPI (Johannesburg), MGUH (Copenhagen), and GSI (Kolkata), but the ones mapped above hold a lot of the world's temnospondyl holotypes and best specimens. Where to find the temnos (Part 2)Now how does this stack up to where temnospondyls are actually found (i.e. where they actually lived)? 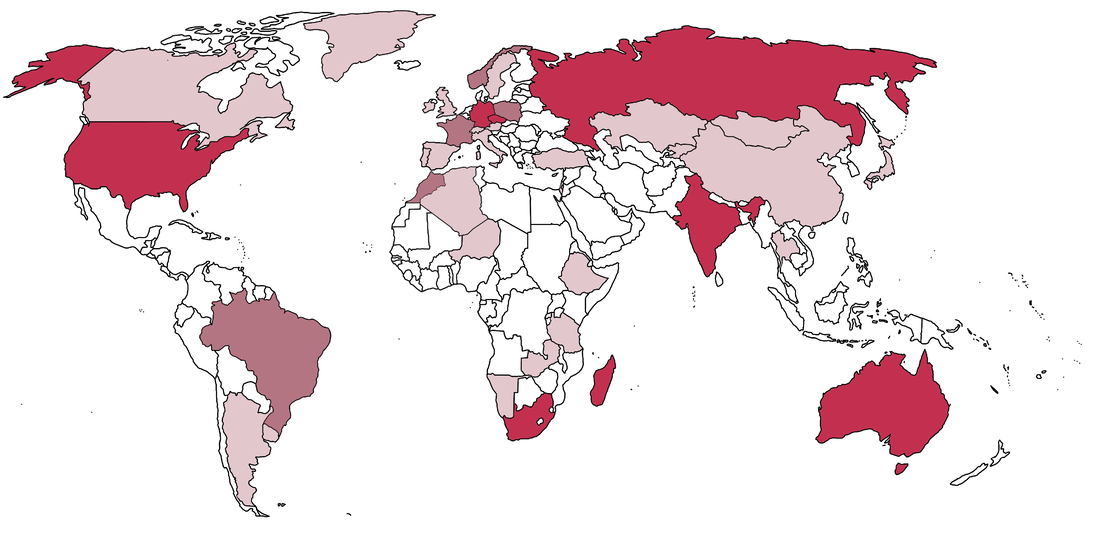 A four-tiered map showing general density of temnospondyl fossils; darker red indicates more fossils, and white indicates none. The lightest red is usually restricted to a single site that has not produced substantial material. The most obvious takeaway is that temnospondyls are all over the world (they're also known from Antarctica, but that's not mapped on here since it's not a sovereign country). If you sort of compare the two maps, there's pretty good alignment between the top temnospondyl-holding institutions and the countries with the most temnospondyl fossils. However, there is one small (geographically speaking) outlier: the U.K. This brings me to the next part of this pseudo-study - are temnospondyls frequently the subject of "helicopter research?" Helicopter scienceThe concept of "helicopter science," where researchers from predominantly western countries in N. America and western Europe figuratively (but sometimes literally) helicopter into an underdeveloped country without comparable research or academic infrastructure, collect fossils, and then publish them without local collaboration in the authorship team, is fairly recognizable today. This can realistically happen in almost any discipline, and there are plenty of good articles out there on the internet (see here, here, and here), but in paleontology, it's particularly common in Africa (excluding South Africa to some degree) and South America. Helicopter research is hardly ahistorical - it is a barely disguised arm of the long history of colonialism, rooted in the exploitation of human and natural resources of underdeveloped regions and populaces, and it remains a huge problem in paleontology because many historical collections were made by European countries in places like Africa and now continue to benefit from the research and public interest valuations of those fossils. Repatriation and other means of reconciliation remain sticky subjects (e.g., Germany's holding of the sauropod Brachiosaurus from Tanzania; source and source). Note that this is different from repatriation of explicitly illegally held fossils, like those smuggled out of Brazil (source) or Mongolia (source); many of the historical collections were made through legal, museum-sponsored expeditions (admittedly at a time when fossil regulations were near non-existent in most countries, perhaps a convenient thing). 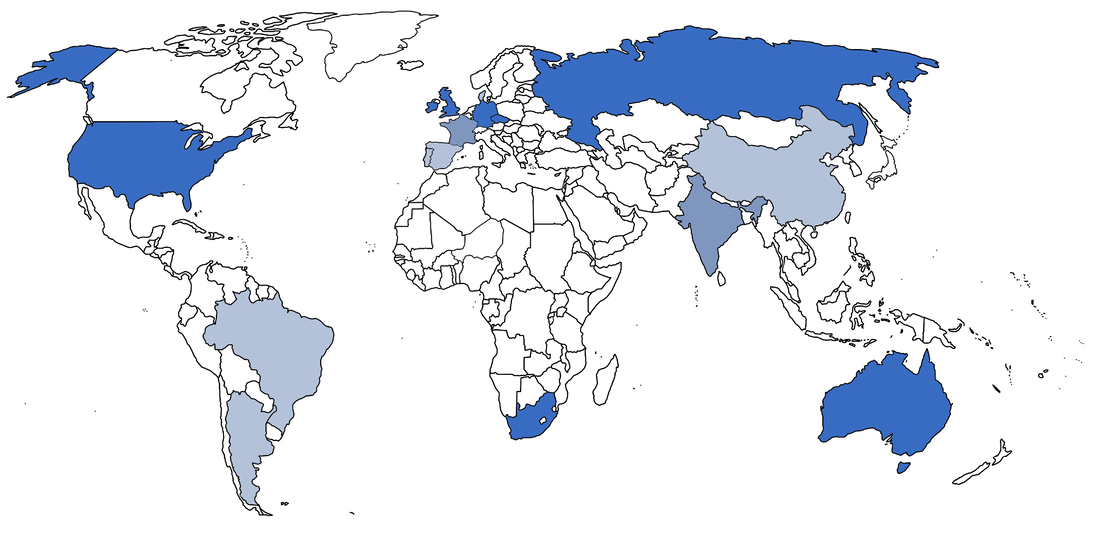 If I put the previous map of major temnospondyl hotspots side-by-side with a tiered map showing major repositories of temnospondyl fossils (in the blues; white only indicates very few to no fossils, not absolutely nothing), the U.K. should pop out as best a small set of islands can. France punches a little above its weight, but nothing like the U.K. So let's talk about the U.K. The sun never sets on the British empireThe U.K. has obviously punched way above its geographic weight in global politics for centuries, and vertebrate paleontology is no exception; other than maybe Germany, it produces more vert paleontology research output than any other European country. Now unsurprisingly, there are not very many fossils in general on the isles; islands are not usually major sources of fossils. But the U.K. holds a remarkable number of not just fossils, but holotypes, from other countries, including a number of ones with fairly well-developed paleontology infrastructure. 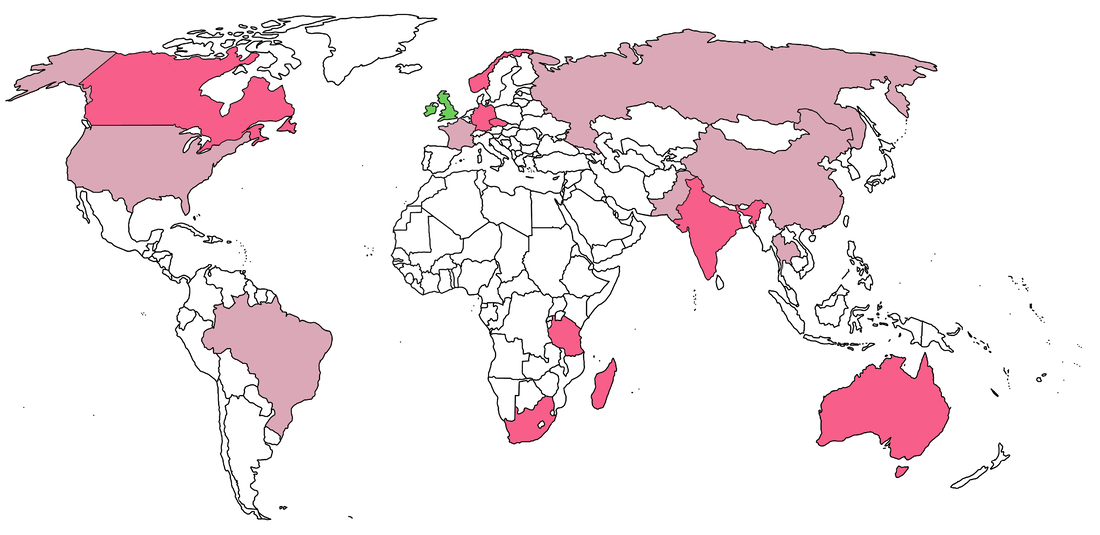 Map showing geography of countries with fossils held in collections in the U.K. The watermelon pink represents specifically holotypes. Note that China and Pakistan are only represented here because of one referred specimen of Kashmirosaurus from the disputed region of Kashmir. Basically all of these exports are housed at the Natural History Museum (NHMUK, formerly the British Museum of Natural History [BMNH]). The list of holotypes housed there is fairly impressive (left) compared to the domestic holdings (right):
Anyone with a fuzzy sense of world history will probably notice that most of these exported holotypes come from countries that were major colonies of the British empire (evidently American independence warded off any prospective future attempts to take most of our fossils). In fact, the only continent that the Brits don't have temnospondyl fossils from is Antarctica, and there are barely any from there to begin with. This changes the discussion a little bit though - the wide-ranging holdings of the U.K. is really just straight-up colonialism rather than helicopter research, which is a term mostly used with respect to contemporary research. Helicopters didn't even exist when most of this material was collected (mainly 18th and 19th centuries). As far as I know, no one in the U.K. is globetrotting to collect more temnospondyls (or has for several decades), although Andrew Milner made an entire career out of working on the extensive holdings of non-domestic, European temnospondyls (especially from the Czech Republic) at the NHMUK. I won't even pretend to know anything about the history of Czech paleontology, but there is a massive amount in the Narodní Muzuem in Prague, so it's not like the country was exclusively pillaged by other countries. What the heck, let's do France too: 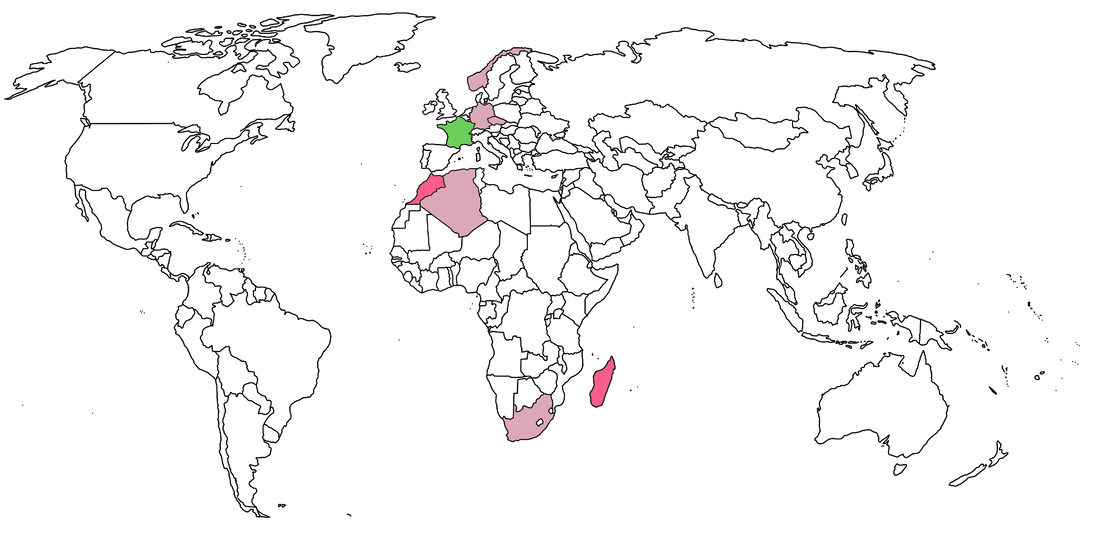 France of course had a much smaller colonial reach by the time that paleontology had taken off as a field. However, they cornered the market on northern Africa (politically and paleontologically), and the MNHN has extensive material from Morocco in particular (four holotypes). The French also outperformed the Brits in Madagascar, holding a number of trematosaur holotypes.
I'll map the U.S. as well, but I'm not even going to begin listing holotypes; there are more than a dozen dissorophid holotypes alone, both from and housed in the U.S. If I had to make a crude estimate, there's probably close to 100 holotypes from the U.S. The main American institutions with foreign holdings are the AMNH and the UCMP. The U.S. probably never developed major forays into other countries with the intent of bringing back fossils since there are already so many in the U.S. I'd be that there are some loan swaps with other countries that aren't described in the literature, but no need to get greedy. South Africa clearly got farmed by literally everyone though. 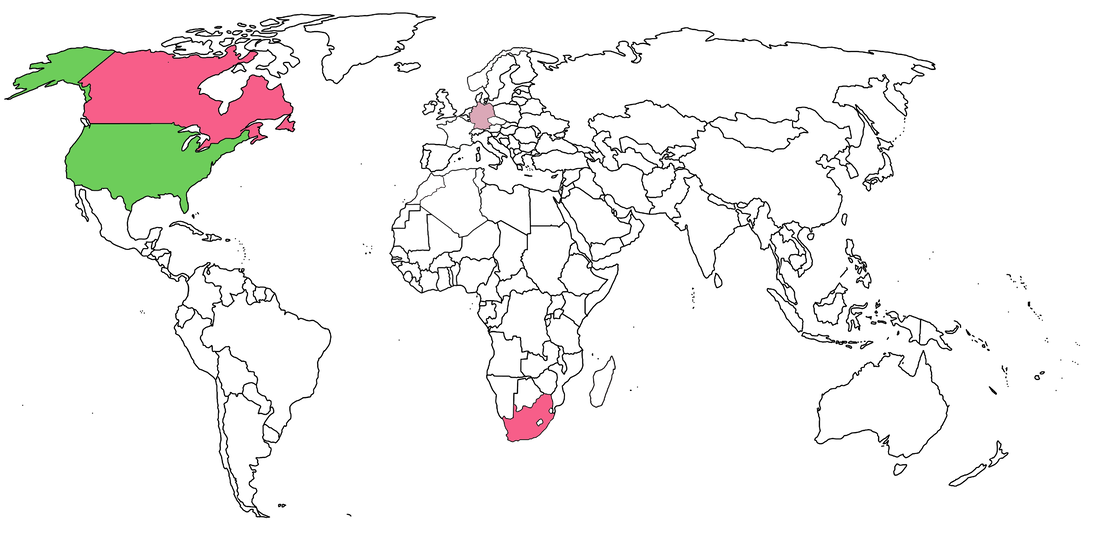 Now in all of this, I want to point out that non-holotype holdings from another country could be swaps - you give me something I don't have, and I'll give you something that you don't have. This happens a lot between museums and is a great way to diversify their holdings and exhibits. For example, there's one Lydekkerina specimen at the Royal Ontario Museum in Toronto, but I'm pretty sure nobody from that museum went down to South Africa just to collect one Lydekkerina... The Natural History Museum in London notes on their website that a bunch of their German material was donated by Herman Credner. Let's come back to the original question of helicopter research. Most of the historical work is direct colonialism, but what about nowadays? To be frank, there isn't that much opportunity simply because nobody's getting a million-dollar grant just to go into a relatively unexplored (paleontologically speaking) region and dig up specifically temnospondyls. Brazil remains largely worked by Brazilians, or at least other Latin Americans (e.g., Eltink et al., 2016; Pacheco et al., 2017; Azevedo et al., 2019; Dias et al., 2020), with minimal involvement from North America or Europe. Asia outside of India has never produced many temnospondyl fossils, most of which are published by local researchers (e.g., Liu, 2016, 2018; Chakravorti & Sengupta, 2018; Mukherjee et al., 2019; Rakshit & Ray, 2020). So Africa is the only other region with the potential for helicopter research, and even that hasn't produced a lot of new material. The track record there is pretty good for local collaboration. There is a single Carboniferous temnospondyl occurrence on the entire continent - the micromelerpetid Branchierpeton saberi from Morocco (Werneburg et al., 2019); the last author on that paper is affiliated with Chouaib Doukkali University (Morocco). Two very fragmentary bones were also reported from the mid-late Permian of Morocco by Steyer & Jalil (2009); Jalil is at the Muséum national d'Histoire naturelle in Paris, one of the major repositories of material from Morocco, but he remains affiliated with Université Cadi Ayyad. South Africa was mostly worked by South Africans historically, and they haven't excavated substantial new material in a while. The only other substantial contemporary study of African temnospondyls is the set of papers reported Nigerpeton and Saharastega from Niger; three of the four papers include local collaborators from Niger (Sidor et al., 2005; Damiani et al., 2006; Steyer et al., 2006), and the fourth is a single-authored festschrift contribution (Sidor, 2013). Perhaps most crucially, the material is reposited at local institutions. A secret life abroadWhat about trafficking of temnospondyls? Laws regarding fossil ownership are famously variable between countries and usually lacking in those without established infrastructure for collections and research. Brazil is the most recent country re-emerge in the news over the dinosaur Ubirajara, a fossil exported to Germany under rather dubious circumstances that suggest a fair bit of impropriety at the bare minimum (Science, NatGeo). Eagle-eyed readers might have noticed that Brazil is one of the countries with specimens housed in the Natural History Museum in London - this material is of Prionosuchus plummeri, the largest known temnospondyl and in fact includes the specific material used to get the approximately 9 m length estimate. The paper describing it, Cox & Hutchinson (1991), indicates the material was collected in collaboration with Llewellyn Price, one of the most famous Brazilian paleontologists, in 1970 and 1972. This falls within the gray area between 1942, when Brazil requires governmental approval for export, and 1990, when the government bans permanent export; obviously at the time, this was not considered an ethical issue, and the paper makes no mention of the export process.
Behind the scenesNow that we've looked at things from a fairly coarse scale, what about the individuals who actually do the research? To get an idea of this, I built out a dataset of publications in two time intervals, 2000-2009 and 2010-2020 (both inclusive). This is mostly grabbed off of Google Scholar, although I got ones that aren't indexed there if I knew about them. Some general guidelines I was using:
Who's whoLet's start by looking at the most productive temnospondyl workers. Note that to keep these graphs somewhat concise (there's several dozen unique first authors from 1999 to 2009 and almost 100 from 2010 to 2020), I'm using an arbitrary minimum cutoff of 6 publications (authorship position irrelevant). I'll mostly show graphics related to this particular subset, but it isn't a referendum on what I think about either publications or authors that aren't captured here because of the cutoff, and it does not mean that everyone who is listed here is a temnospondyl "expert," either as self-defined or as perceived by others. That being said, many of the names on here are the first people that the average paleontologist would think of for temno-related matters. The point of these graphs is just to show numerical production. Citation count or other measures of impact take longer to data-mine and get complicated when you can't use a single search engine to get measures for every study, not to mention that 11-year sampling bins are not as good. 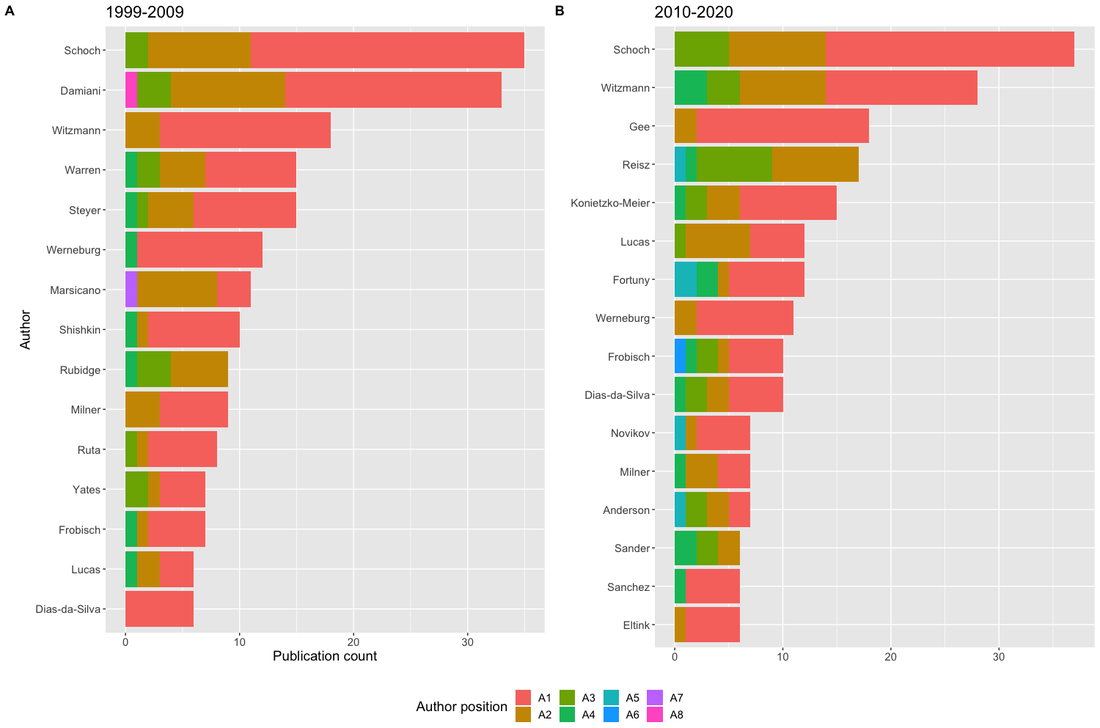 I think the plots are pretty self-explanatory, and they show some interesting patterns. Firstly, two German dudes, Rainer Schoch and Florian Witzmann, have dominated the temnospondyl literature (note that their true net contribution is overestimated by these plots because they frequently co-author papers, at least 10 on a quick count). Country trends will be discussed next, so I won't get into them too much here, but there's a few other areas to comment on that aren't clear from the plots themselves.
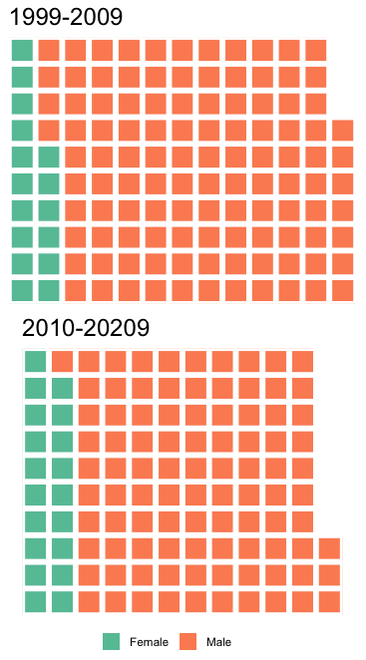
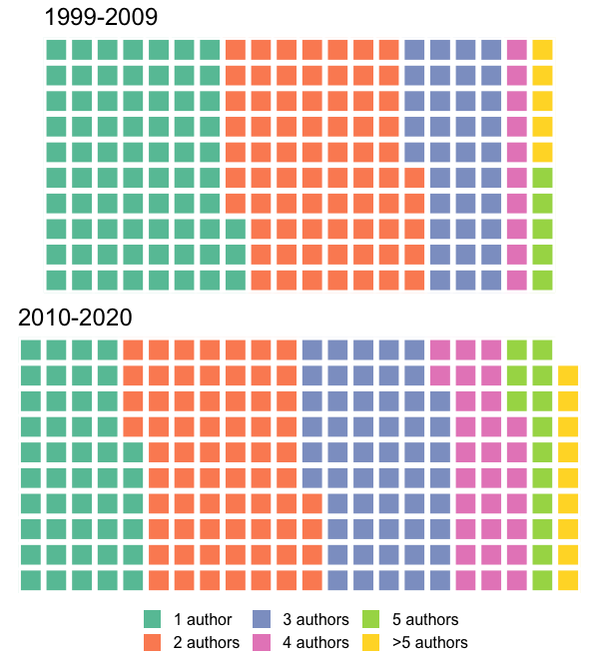
As an aside, it might look weird that the 1999-2009 interval has more publications, but what this really reflects is a declining "market share" of the entire set of publications by this arbitrarily defined subset that I'm using here. Both of these subsets represent a majority of the total output of each time interval, but it's much more substantial for the 1999-2009 interval (63.5% for 1999-2009; 52.1% for 2010-2020).Between 1999 and 2009, there were only 66 unique first authors; that number went up by essentially 50% to 97 unique first authors between 2010 and 2020, and as a result, a smaller proportion of the total papers were produced by the subset of the most productive workers. Whether this increase in distinct workers reflects increased interest or just increased access can't be addressed here. Now let's back out and check the entire dataset - is it possible that women are overrepresented below the cutoff of >5 publications that I defined here (i.e. that women are either less likely to be first authors or that they produce fewer first-authored publications than men; not mutually exclusive)? Indeed, the proportion of women as first-authors is higher in the overall dataset for both time intervals (17.3% and 25.2%, respectively) than in the subset of the most productive workers (12.6% and 16.8%). 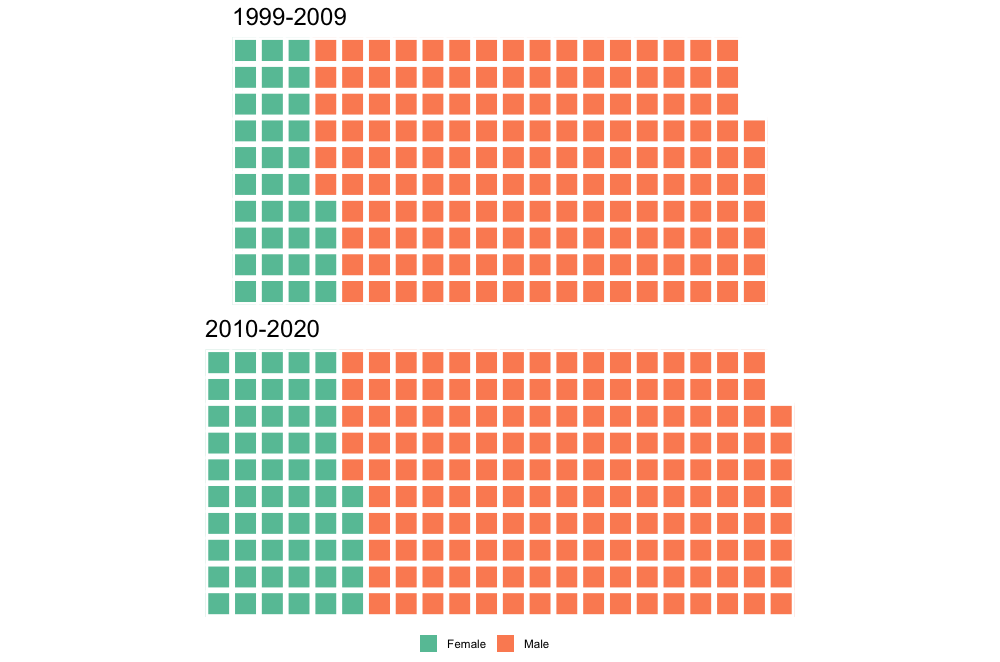 Indeed, between 1999 and 2009, there were 16 unique women who first-authored a paper, but only three of those published more than five papers in that time (women were 24.2% of all unique first-authors); between 2010 and 2020, there were 28 unique women first-authoring a paper, and also only three of those published more than five papers in that time (women were 28.8% of unique first-authors). By country of affiliation of lead authorNow let's back out and take a look at how things break down by where the primary listed affiliation of the first author is physically based for all of these studies. 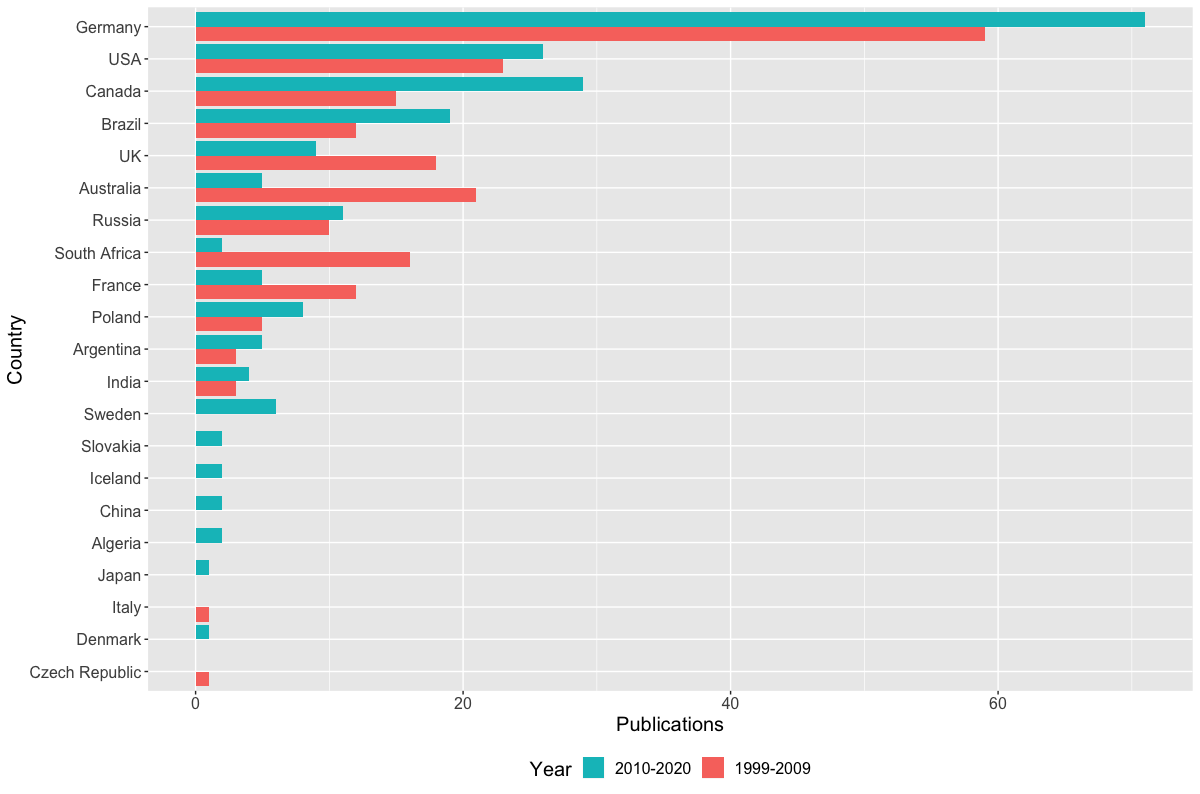
'The single genius'Science is often fixated on this concept of single geniuses (coincidentally mostly men) where we elevate these really well-known figures like Einstein, Hawking, Darwin, Newton, etc. as incredible geniuses who essentially accomplished their major findings on their own. This has drawn a lot of criticism in recent years because it often totally obfuscates the actual history of discovery, whether in Crick & Watson's overshadowing of Rosalind Franklin, or in the more everyday aspects of scientific research where PIs claim credit for student work. Some reads here (BBC), here (New Republic), and here (Vox). People don't have to be scientists or Nobel laureates to have this title figuratively bestowed upon them (or to place it upon themselves); we are all cultured to think of the people at the top as the 'masterminds' or the 'big brains' behind a complex machine, whether in film (directors, producers) or in science (PIs).
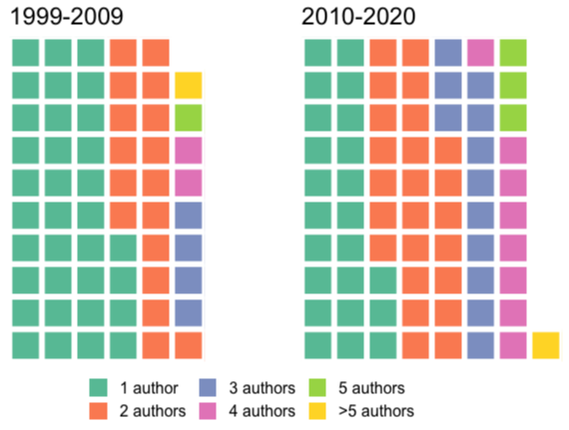
The TLDR is that there is a definite trend toward low authorship teams, though I obviously don't have comparative data to assess how that stacks up against other groups of tetrapods. Now there's plenty of ways to speculate on why temnospondyl research is skewed towards small research teams. One, of course, is that certain workers do perceive themselves in this 'genius' vein and either feel that additional collaborators have nothing to contribute or will in fact slow them down. That's sort of pessimistic, but it's inherently true that there is only one limiting actor in a single-authored project, which could be good or bad depending on how you work and what you're working on. Re: the latter, there might be less of a perceived need for collaboration in descriptive papers, which remain the bulk of temnospondyl analyses - all you need is one well-traveled person who has seen a lot of the relevant material in theory (this is not a personal advocacy for avoiding collaboration in descriptive papers). A third factor is that some of the most productive workers are based at museums with good temnospondyl holdings (e.g., Fröbisch, Lucas, Schoch, Steyer, Witzmann) - that means that not only may they not have to go anywhere to look at specimens, there's also a smaller chance that authorship associated with another curator granting access to collections is involved. One other possibility is trying to avoid COI (conflict of interest) for a certain period of time. I don't know how it works in Europe, but here in North America, frequent collaborators are usually banned from reviewing your grants ('frequent' for NSF is defined as any collaboration within the last year, graduate supervision, and a few other activities). The same thing was a problem for me when trying to find an external examiner for my PhD defence (most universities require that you have someone not affiliated with the university) - Toronto requires someone who has themselves graduated a PhD student and who has not collaborated with you or your advisor in the last 5 years! I can certainly appreciate that it is already hard enough getting temnospondyl reviewers for papers (which have far looser COI rules, which are really more like "suggestions"), and I think that this may also influence how people go about forming research teams. If I had to guess though, the main reason why temnospondyl teams are small is because the temnospondyl community is also small. To pass muster, you gotta find at at least two and maybe three or four reviewers to green light the paper. That's a lot (I have problems getting more than two a lot of the time), so any potential collaborator is also a potential reviewer - put them on the paper and now you've knocked down your reviewer pool. I have definitely had non-temnospondyl workers review some of my papers, and I don't think that's they were the top choices either. Mathematically, needing at least two reviewers per paper means some people get asked to review a lot, and for any given reason, someone might not be able to do it. You might also just really like somebody as a reviewer - I have certain people who I find to be very good reviewers (=/= short review necessarily) - so regardless of whether that person is also a great scientist, you might leave them off the team unless you think that they are essential (hard to argue for essentiality in most descriptive work). TLDR
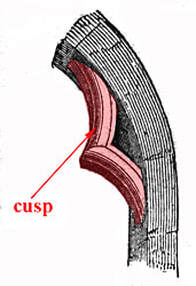
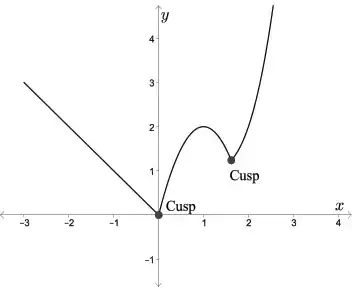
The average person is probably most accustomed to thinking of 'cusp' along the lines of "on the edge" or "on the verge." As in, "we're on the cusp of returning to a normal life with this vaccine." But 'cusp' actually has a different simple meaning, which is derived from the Latin word for 'point' or 'apex.' Architects use it to refer to a feature that became particularly popular in the Gothic period, a small projection between small arches. Mathematicians use it to refer to the point where a curve drastically changes directions. And biologists use it to refer to either a tissue fold that contributes to a valve to prevent blood backflow or to a pointed protrusion on the surface of the tooth. (There's also usage in astrology, but I'm not getting into that). I'm not an architect or a mathematician, nor do I work on blood vessels, so here we are talking about teeth!
A decidedly non-model systemIn the old days, a lot of what we inferred about other "complex" animals was based on what we knew about humans, since we knew ourselves best (hypothetically), which is why a lot of anatomical features were first named based on humans. Of course, if you go to a nice nature area and look around, it's pretty easy to see that humans are not really a great "model" for other animals beyond primates. Ignoring things like complex societies and technology, we walk on two legs but can't fly, have reduced hair, and have opposable thumbs, features that are mostly shared (through common ancestry) only with other primates. Mammals are generally bad models for other animals, which is why when scientists refer to "model organisms" that are used for their combination of adaptability to lab settings and applicability to a wide range of biological systems, we don't use many mammals. Anyone who's been through a high school or college biology class probably knows a little bit about the humble common fruit fly (Drosophila melanogaster), or perhaps the microscopic nematode Caenorhabditis elegans (usually just called 'C elegans' to dodge the wordy genus name), or maybe even the quietly invasive African clawed frog (usually called Xenopus by scientists and not to be confused with the African dwarf frog that Petsmart sells). These are considered model organisms for many reasons, none the least of which is that they're very hardy in lab settings, but also because their anatomy/physiology makes them reasonable proxies for something that's more complex and harder (or impossible) to work with in labs. At least some readers of the blog may be surprised to know that in the 50's, the frog I mentioned was used as a living pregnancy test for women with shocking accuracy (details in article; frogs not really harmed in process). I'm pretty sure it would be illegal to do the test in reverse to test whether a frog was pregnant by experimenting on a human. Which is one reason why Xenopus continues to be a model organism despite being discontinued as a pregnancy test.
Coincidentally, Xenopus is also a good model for the topic of today's blog post - the primitive tooth condition in tetrapods. Teeth come in a wide variety of shapes, none more evident than in our own mouth, in which the various tooth types are named as such because they look different. Most animals don't have this kind of differentiation though (called 'heterodonty'); we call animals without differentiated teeth 'homodont.' Look in the mouth of the fish on your cutting board, and you're likely to see a lot of teeth that all look the same. They probably look quite simple too - simple cones with a single point. We call this tooth 'monocuspid' because it has one cusp. Teeth with two cusps are 'bicuspid,' teeth with three cusps are 'tricuspid,' and so on (though usually 4+ is just called 'multicuspid'). Monocuspid teeth are found in Xenopus, as well as most early tetrapods, including most temnospondyls.
But Xenopus is an outlier among its froggy friends. Most frogs actually have bicuspid teeth (although toads, which are frogs, have no teeth), and a handful are reported to have tricuspid teeth (e.g., Greven & Ritz, 2009), but none have more than three. If you want an example of a tooth with more than three cusps, just look at your own molars - there are 4-5 in humans. Bicuspid teeth are typical for salamanders and caecilians, the other two groups of living amphibians, but they are not super common in animals with homodont dentition or are only found in part of the mouth (our premolars are bicuspid, which you maybe heard your dentist say when looking at your x-rays). Amphibians usually have homodont dentition (same size and shape throughout the mouth), so naturally, this has put an emphasis on searching for bicuspid teeth in the fossil record as one means of elucidating the origins of modern amphibians. 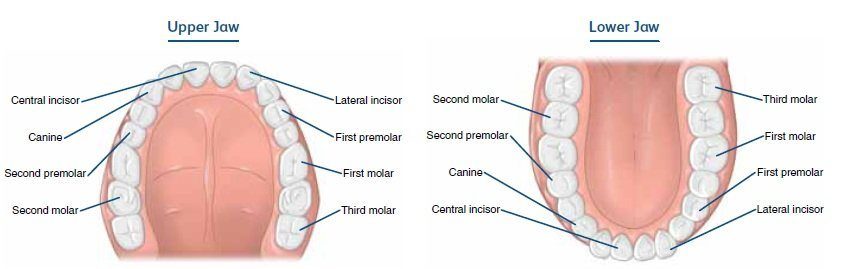 Diagram showing different tooth types in the human mouth. There are two premolars on both upper and lower jaws that are also called 'biscuspids' for their two cusps. Image credit: ACI Medical & Dental School. Tales of the teethLike I mentioned before, monocuspid teeth are the base model for tetrapods and temnospondyls. This isn't really surprising; adding points is complex and requires more nuanced spatial control over the tooth's development rather than just forming one point. There is only one example of a temnospondyl with tricuspid teeth, the branchiosaurid Tungussogyrinus from Siberia (Werneburg, 2009). The broad acquisition of more complex teeth, as well as heterodont dentition, tracks the expansion of tetrapods into different ecological roles, more specifically their dietary preferences. Scraping algae off of rocks, for example, is apparently a key driver of tricuspid teeth in marine iguanas (e.g., Melstrom, 2017); this idea has been extended down to some extinct tetrapods with tricuspid teeth as well (e.g., Mann & Maddin, 2019). Temnospondyls, like amphibians today, aren't picky, but they aren't generalists either. Amphibians can't really eat plant matter, for example, whereas the capacity for herbivory was an acquisition that facilitates the diversification of amniotes on land. But temnospondyls did eat other animals, and for that, monocuspid teeth work just fine, whether for grabbing, piercing, or stabbing.
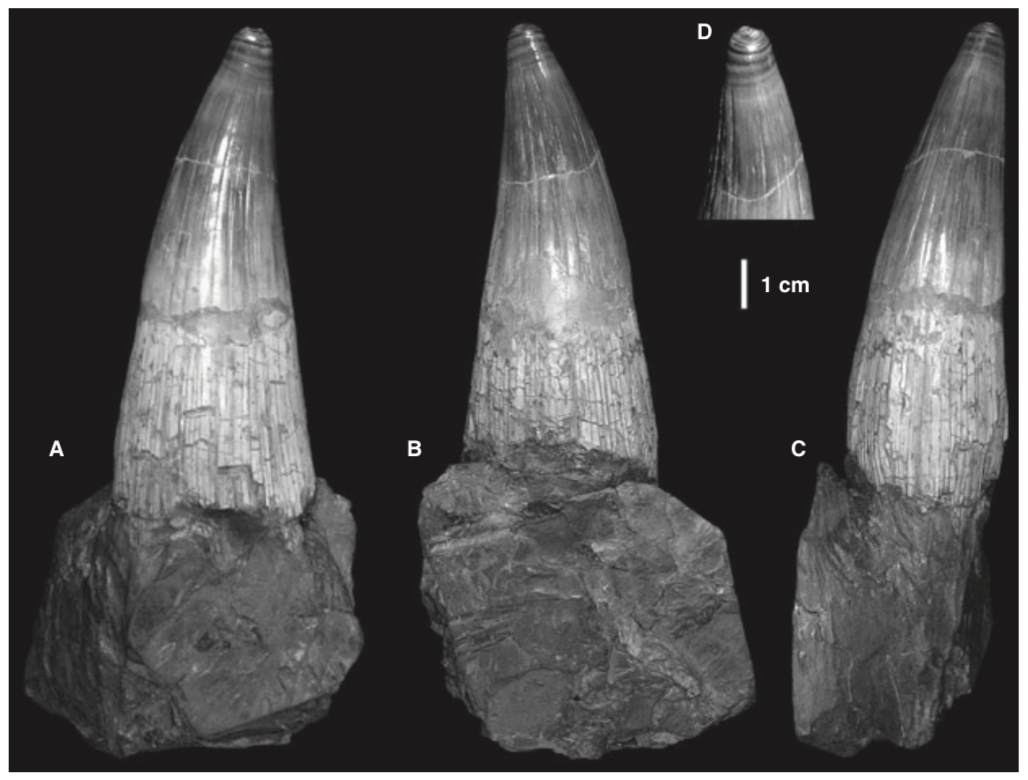
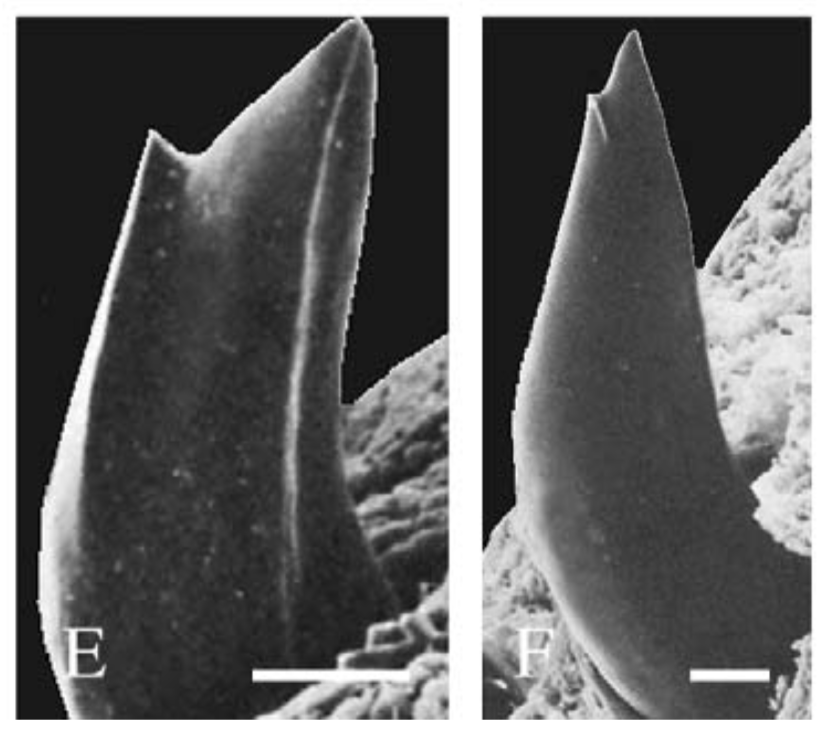
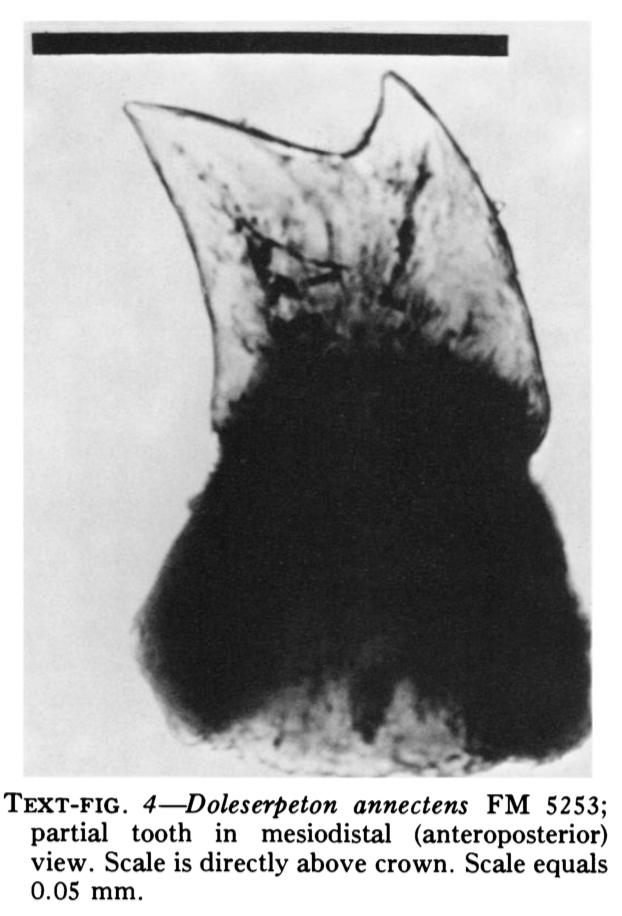
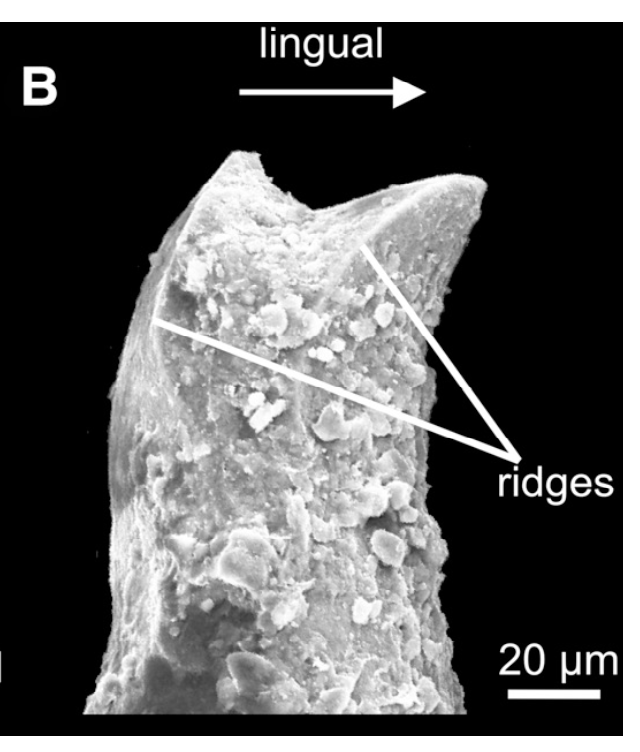
There are several examples of temnospondyls with bicuspid teeth, or to be a little safer, several taxa in which bicuspid teeth are reported. Probably the most unequivocal of them is Doleserpeton annectens, which John Bolt keyed in on in the 1960s as a "proto-lissamphibian" based on his observation of this feature. With a lot of material and excellent preservation, subsequent research has confirmed the presence of bicuspid teeth in this taxon (e.g., Sigurdsen & Bolt, 2010).
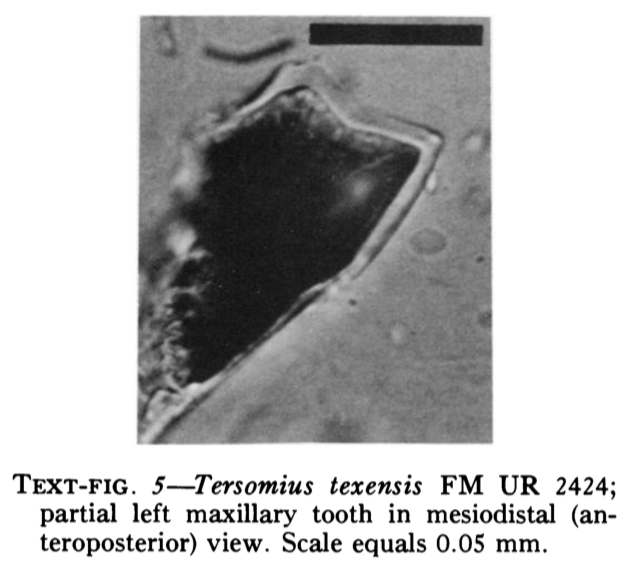
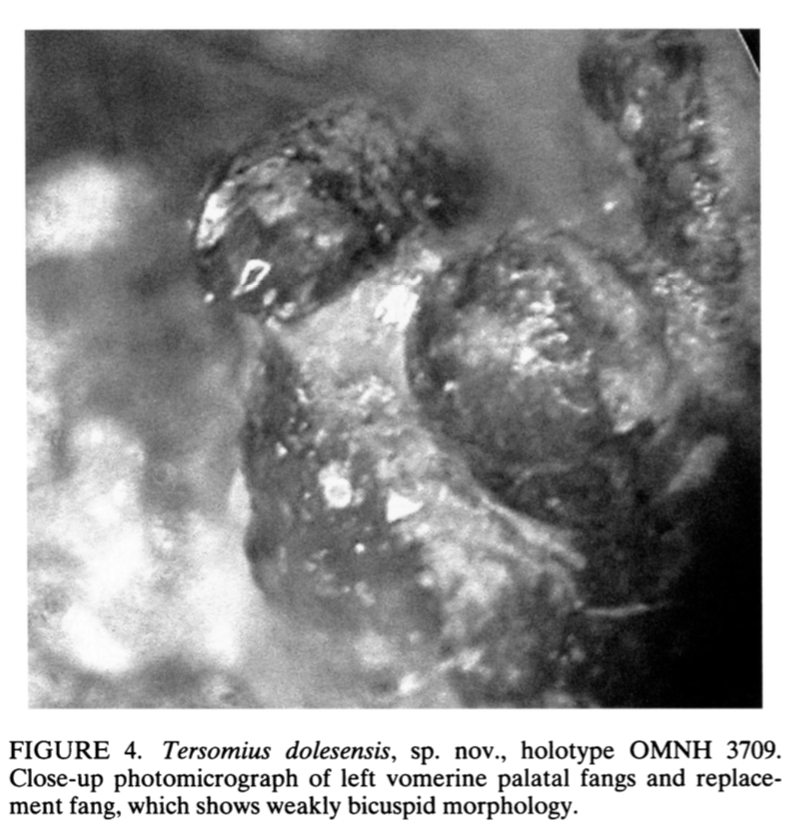
With the recognition of this feature in one amphibamiform, workers began looking for more evidence of bicuspid teeth in what was then called 'amphibamids.' Amphibamus, a close relative of Doleserpeton, also has bicuspid teeth (Bolt, 1979). Platyrhinops, which is somewhat debated over whether it is the same as Amphibamus, also has bicuspid teeth (Clack & Milner, 2009). Most amphibamiforms do not have bicuspid teeth however, including Gerobatrachus, the 'frogamanader' ('batrachian' is the technical term for the grouping of frogs and salamanders; Anderson et al., 2008). There is also some controversy about whether any species of Tersomius has bicuspid teeth. Bolt (1977) indicated some specimens of Tersomius texensis (but not the holotype) have bicuspid teeth, but he only figured one of these bicuspid teeth (it was hard to get clear photographs or microphotographs in the 70's). The specimen that he did figure does appear to have two cusps, but they are not nearly as well-developed as in Doleserpeton or in most living amphibians. Probably important to bear in mind here is that we're talking about teeth that are less than one-tenth of a millimeter in width, so the precision in the plane of sectioning could be a factor. Notable too is that all of the Tersomius material with bicuspid teeth is from the South Grandfield site in Oklahoma; possibly, it is not actually T. texensis (or Tersomius at all). The original assignment was provisional (cf. designation by Daly, 1973) and made no comments on the teeth. That material has not been redescribed. This is further complicated by Anderson & Bolt's (2013) description of Tersomius dolesensis. Not only does it have the peculiar combination of monocuspid marginal teeth with at least one bicuspid palatal tooth, but there is disagreement about whether this is actually properly placed in Tersomius (their phylogenetic analysis excludes other species). In Maddin et al.'s (2013) analysis with the type and one good referred specimen of T. texensis, T. dolesensis does not group with what are now termed micropholids (Pasawioops + Micropholis + Tersomius). Suffice it to say that some amphibamiforms have bicuspid teeth and most do not.
 Monocuspid teeth of the amphibamiform Gerobatrachus (Anderson et al., 2008) - this is one of the closest relatives of at least frogs + salamanders under many hypotheses. Monocuspid teeth of the amphibamiform Gerobatrachus (Anderson et al., 2008) - this is one of the closest relatives of at least frogs + salamanders under many hypotheses. Unsurprisingly, paleontologists have looked to other possible lissamphibian candidates, primarily 'microsaurs,' to see whether they too might have bicuspid teeth (diminishing the value of bicuspidity in amphibamiforms). And indeed, there are numerous reports of bicuspidity among 'microsaurs,' including Carrolla craddocki (Langston & Olson, 1986; Maddin et al., 2011), the now defunct 'Bolterpeton carrolli' (Anderson & Reisz, 2003; Bolterpeton is synonymous with the parareptile Delorhynchus), and Cardiocephalus peabodyi (Anderson & Reisz). The description of two cusps in C. craddocki still stands, whereas that of C. peabodyi seems to have been quietly abandoned. Haridy et al. (2017) suggest that in taxa with cutting edges on the teeth, like in many parareptiles, these edges may be misinterpreted as the ridges leaving to two cusps; they specifically suggest that what used to be 'B. carrolli' appeared to have two cusps only because of uneven tooth wear. It's also worth noting that the putative stem caecilian Chinlestegophis, a stereospondyl temnospondyl, also lacks bicuspid teeth (Pardo et al., 2017), as does the more highly nested amphibamiform Gerobatrachus (Anderson et al., 2008). It's just a phaseOne of the key caveats in bicuspidity in modern amphibians is that it is known to change in the development of the animal. Teeth are monocuspid early in development and only later do they become bicuspid. This is one of the reasons why John Bolt proposed that some amphibamiforms might in fact be juvenile dissorophids, and he proposed the inverse scenario in which the adult stage was a monocuspid tooth that transitioned from a bicuspid tooth. 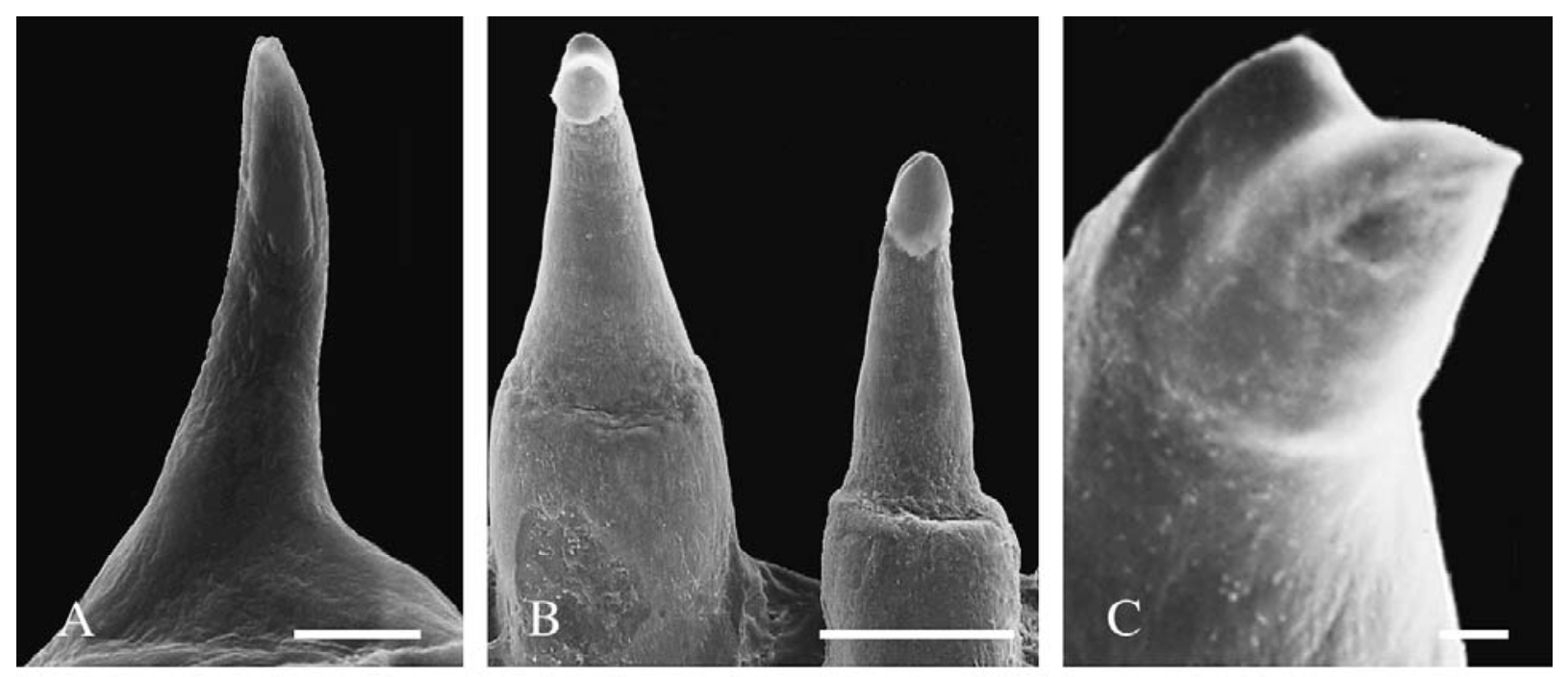 Comparison of teeth of the Iberian Ribbed Newt at the larval (A), juvenile (B), and adult (C) stages. Figure from Davit-Béal et al. (2007). Features that are transient throughout an animal's life become difficult to work with in the fossil record because it means that absence (of bicuspidity in this case) is not necessarily attributable to a legitimate absence of homologous features - instead, the material under study might be immature and not yet have achieved the derived condition that is being sought but would have at a more advanced stage of life. Bolt wasn't the first to suggest that some terrestrial amphibamiforms might actually be juveniles of larger animals (same problem with the mostly aquatic branchiosaurids), but the data collected since his work in the 70s and 80s has not supported this conjecture. A higher purpose
References
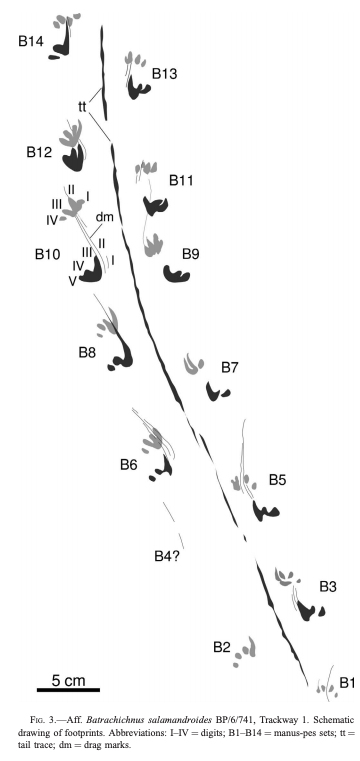
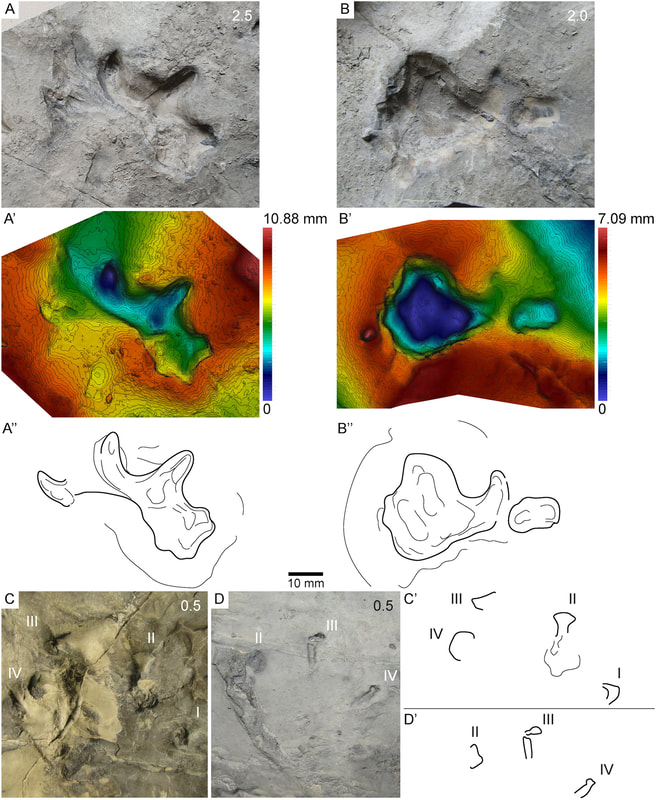
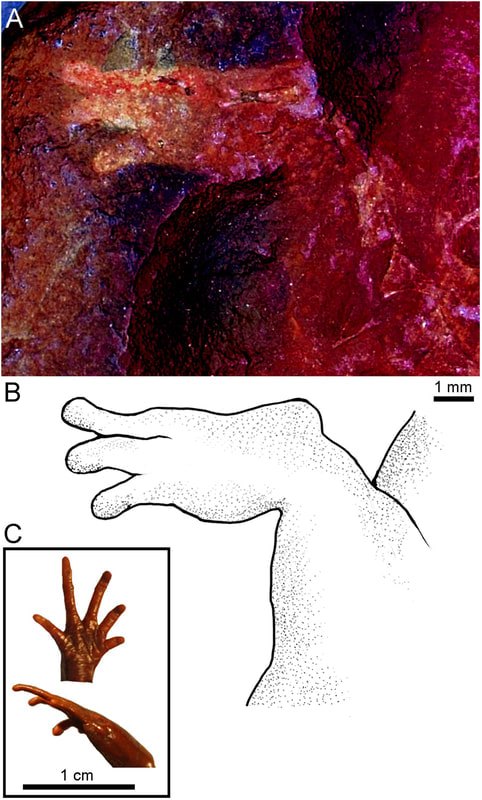
2020 has certainly been some kind of year, which makes it easy to miss the latest scientific discoveries related to everyone's favourite (usually) four-fingered tetrapods. Last year I did a wrap on the entire decade, so there was a quite a lot, but in spite of the circumstances, 2020 has still produced some very exciting temnospondyl research! As with last year's wrap-up, this focuses on temnospondyl-centric research; there are obviously plenty of papers that make cursory mention of them or that might include a picture or two in an assemblage description, but those are not summarized here (I gotta be efficient with my time). As usual, links to everything are in the reference list as the end. Hopefully looking forward to getting back on a more regular track again in 2021 - stay tuned! Leaving a markIn lieu of body fossils, we often have evidence of temnospondyls in the form of trace fossils, which usually consist of a trackway. In contrast to amniotes, which basically all have pentadactyl (5-digit) hands, temnospondyl trackways have been considered to be historically easy to ID because of the tetradactyl (4-digit) hand that typifies temnospondyls and many modern amphibians (at least the ones that still have hands). Body and trace fossils rarely mix because the optimal conditions to preserve both the hard parts and the traces of activity will differ somewhat starkly, but this is good because it means where we might lack a body fossil record, perhaps we could supplement the total record with traces.
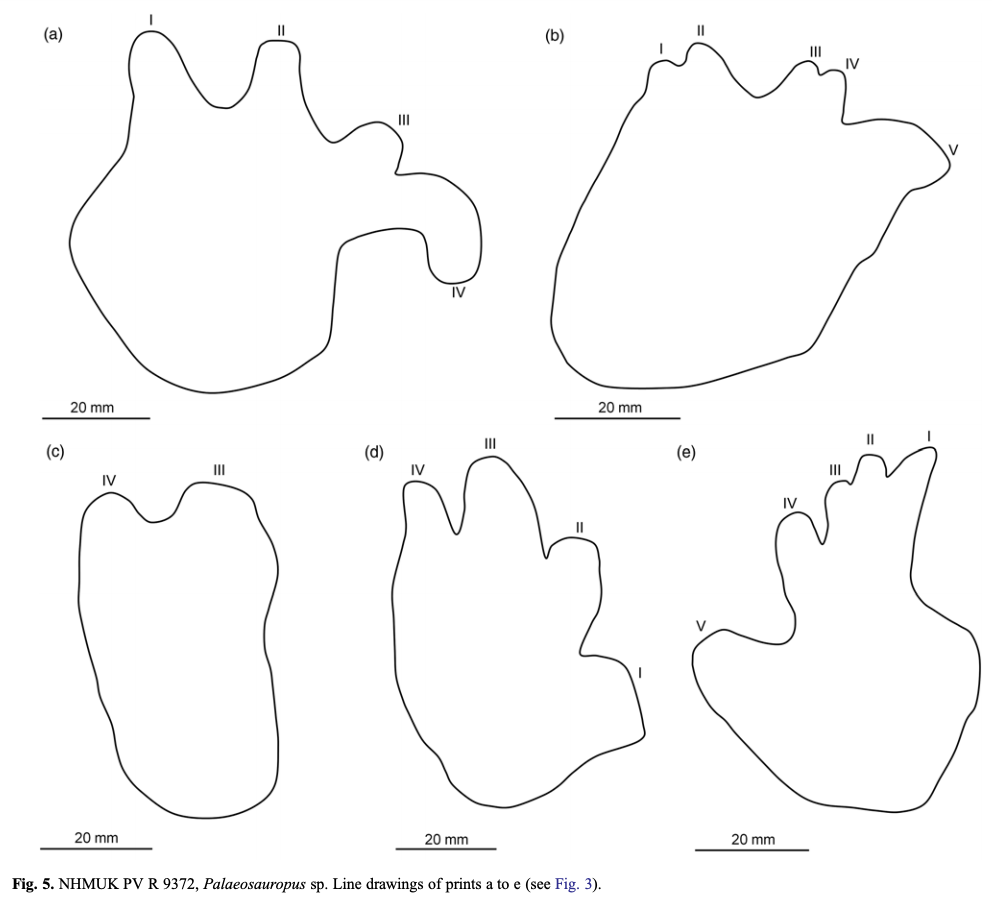
Bird et al. (J Geol Soc) reported a trackway from the early Carboniferous (Visean) of the U.K. (paper technically online in late December last year, but in print this year). This time period is a critical window of tetrapod evolution, when stem tetrapods really radiate; however, we still don't have most of the groups that directly lead to modern groups, and there is only one Visean temnospondyl, Balanerpeton woodi from Scotland (which is also debated about whether it might be a stem tetrapod and not a temnospondyl). The U.K. in general is pretty sparse on the Paleozoic tetrapods. As such, this is a good example where the trace fossil record pre-dates the main body fossil record and thus hints at a large gap in the body fossil record of the earliest stages of a group's evolution (temnospondyls in this case). The ichnotaxon (taxonomic framework for trace fossils) that they assigned the material to is Palaeosauropus, which is also known from the Blue Beach locality in Nova Scotia and Pennsylvania and which was previously interpreted to belong to an edopoid (the validity of this association is a little questionable). The oldest body fossil record of an edopoid is the Bashkirian, which begins around 323 million years ago, whereas the Visean ends at 330 million years, so at minimum, this possible edopoid record extends the group back by at least 7 million years, which is congruent with phylogenetic analyses that recover edopoids as a very early diverging group.
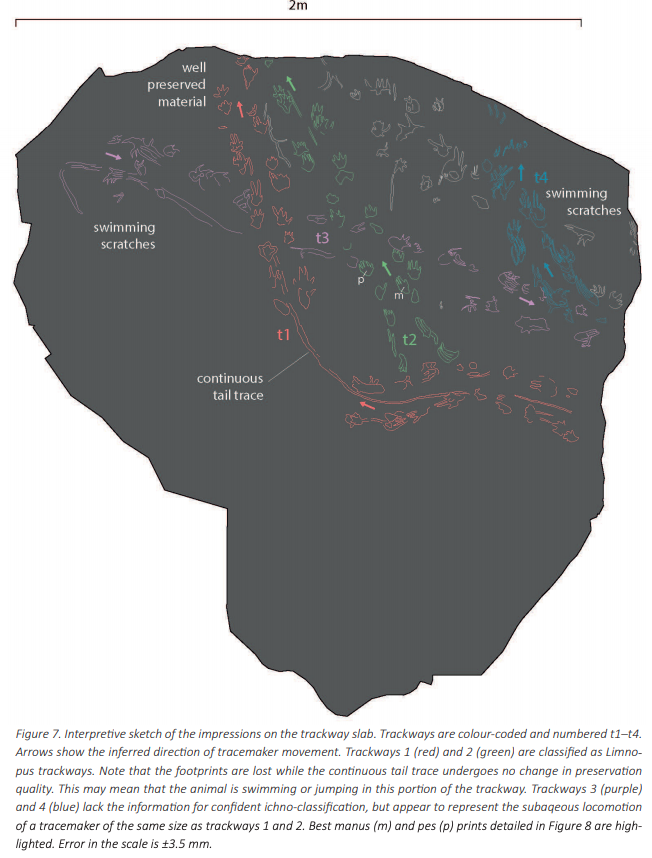
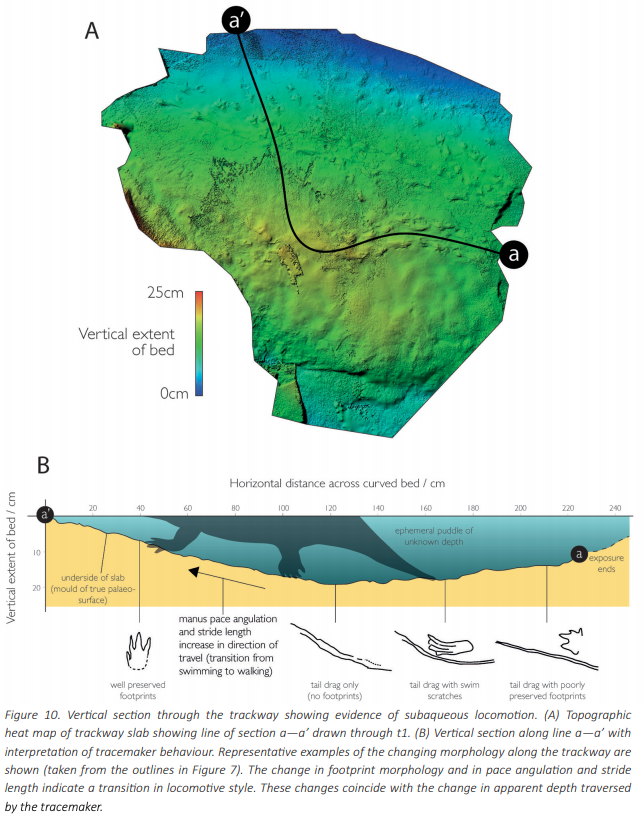
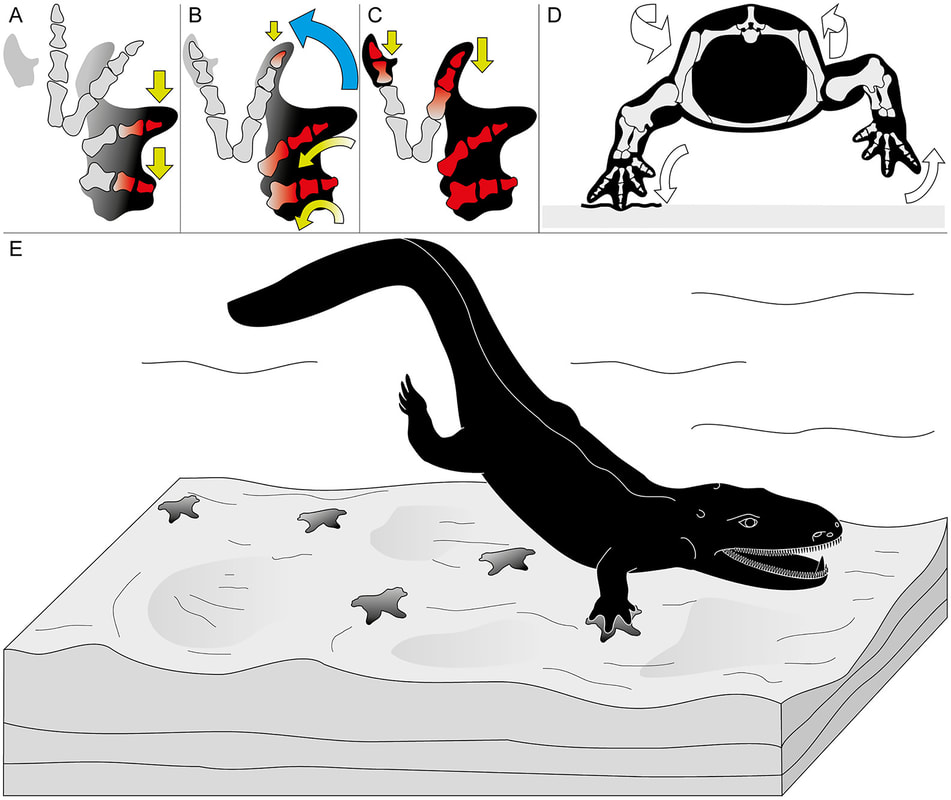
Herron et al. (Norwegian J Geol) reported slightly younger tracks from the Late Carboniferous (Moscovian-Kasimovian), when the temnospondyl body fossil record is much better established, from Svalbard. This archipelago is usually better known for its Triassic temnospondyl body fossil, which compares favourably with the closely situated Greenland assemblage, and the Scandinavian Arctic region has long been productive for some of the most important stem tetrapod fossils like Acanthostega. The new material that Herron et al. report is a massive block (almost 800 photos to produce a composite model) that captures a rare setting: the margin of a pond. As a result, the trackway captures the transition between the animal moving in the deeper part of the pond (with more scratch-like traces made through minimal contact with the pond bottom) and then moving out onto land (with more pronounced footprints).
There were a few other papers that briefly show some temnospondyl traces or trackways that might be temnospondyls:
An abundance of skepticism
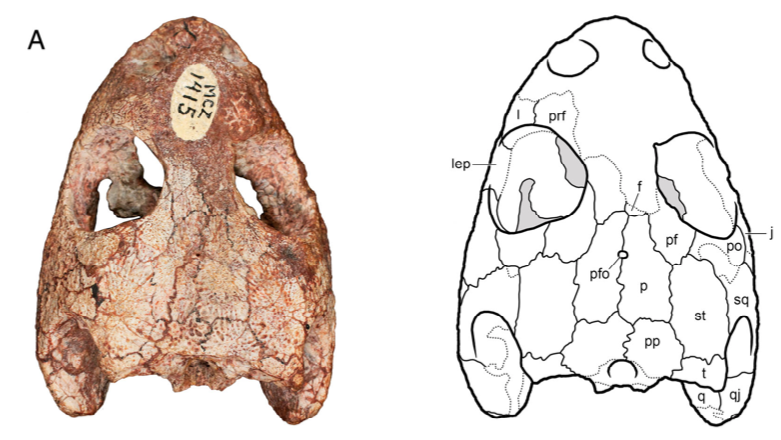
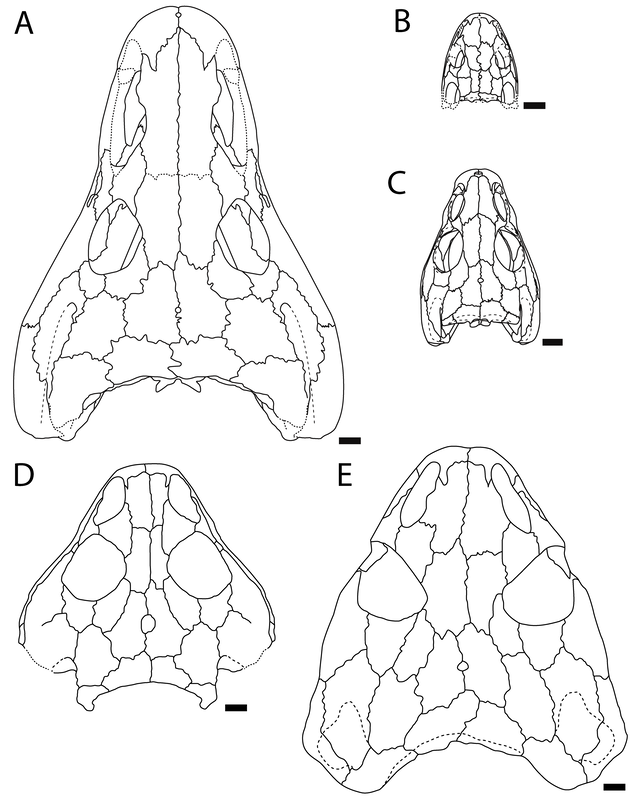
 The second study was a broader review of the fossil record of caecilians by Santos et al. (Biol J Linn Soc); the second author (Michel Laurin) is a longstanding proponent of a single origin of lissamphibians from lepospondyls. This paper repeatedly calls out various lines of argument used by Pardo et al. and even has a discussion section titled "Chinlestegophis: a true gymnophionomorphan?" In fact, they go so far as to say that every identified synapomorphy is questionable, and other shared features are plesiomorphies found in other temnospondyls. The issue of whether the element at the front of the eye is a lacrimal or a LEP is just one of those questioned features. No new phylogenetic analysis here, but a great summary of the available information on fossil caecilians (it is very poor) like the figure below on the left. In case you're wondering why the record is so bad, burrowing animals by virtue of where they live are not nearly as likely to end up near the types of environments that most frequently preserve in the fossil record, like ponds or floodplains. Their small size, like that of other lissamphibians, is another issue that is generally less favourable for preservation.
The dissorophid dynastyThe temnospondyls long considered to be closely related to lissamphibians in one form or another, dissorophoids, continued to see major research interest, even outside of the context of lissamphibian origins. 2020 mostly brought re-descriptions, though we got a few other types of studies thrown in there as well.
One important redescription that we got in 2020 was of the holotype of "Dissorophus" angusta, a species named by Bob Carroll in 1964. Basically everybody has recognized for decades that it is definitely not Dissorophus, lacking the large first osteoderm and having narrow osteoderms throughout the body, but it is only now, 56 years later, that its status is finally resolved as a new genus, Diploseira, by Dilkes (J Vert Paleontol). The only known specimen is mostly the postcranial skeleton, which makes it hard to compare with many other dissorophids, but it definitely has distinctive features. David's usual impeccable attention to detail and illustrations really flush out every little nuance of the anatomy. 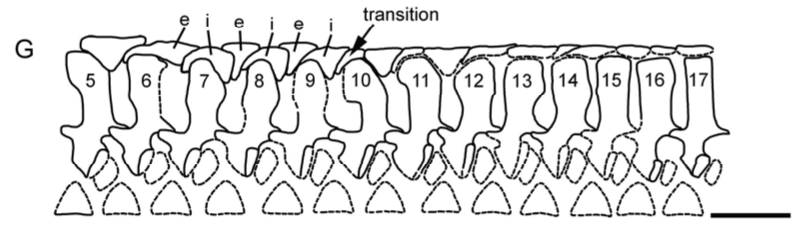 This taxon is quite interesting in preserving a transitional series of osteoderms; there are two series at the front and only one at the back. As far as we know, this is the only dissorophid with this condition - other species either have a single continuous series or two continuous series (external and internal). Much of the osteoderm anatomy (though not the width) is shared with dissorophines (Broiliellus and Dissorophus), and Dilkes' phylogenetic analysis indeed recovered Diploseira within that group. This really mucks up how we use qualitative patterns of osteoderms to make taxonomic frameworks, continuing to highlight the issues with single-feature phenetic taxonomy (essentially arbitrary emphasis of certain observational data).
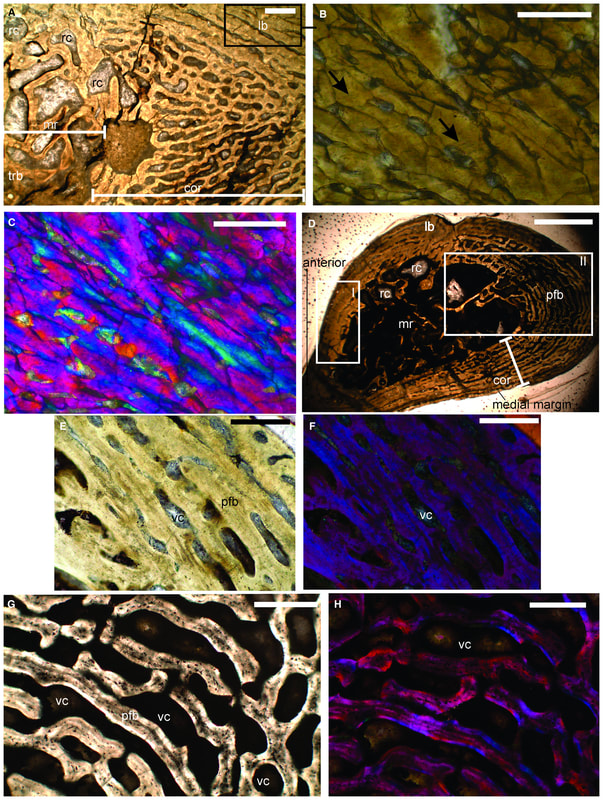
A slice of lifeIn addition to my Doleserpeton paper, there were a few other studies that sliced and diced up some temno bones. The first one was technically published online last year (in print this year), and I very briefly mentioned it in my year-end summary last year, but it's worth discussing here in greater detail. Mukherjee et al. (Papers Palaeontol) describe the histology of a few Middle Triassic capitosaurs from India, which is a follow-up to another paper led by Muhkerjee a decade ago on a more preliminary sample of Indian temnos. Despite the material of the two taxa (Cherninia and Paracyclotosaurus) coming from the same small locality, they show distinctive differences in their bone histology. Cherninia shows a lot of what temnospondyl workers term 'incipient fibrolamellar bone,' which generally meets the criteria of fibrolamellar bone in amniotes. Note that contrary to some oversimplifications, fibrolamellar bone is not an unequivocal hallmark of endothermy - it only indicates rapid growth, which can be accomplished in ectotherms living in harsh conditions that require fast growth (for example, taxa with larval forms that need to get out of water before it dries up).
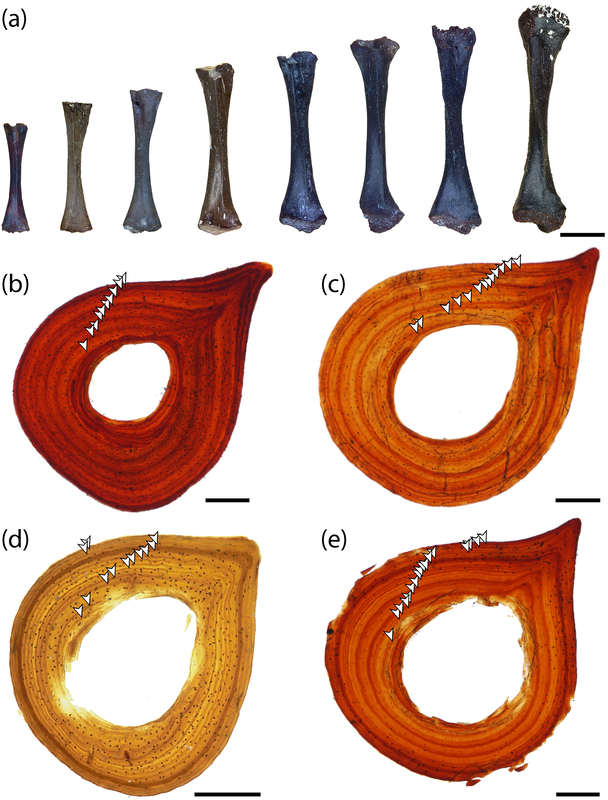
Comparison of histology of the humerus of Cherninia (on the left) and Paracyclotosaurus (on the right). In Cherninia, this fibrolamellar bone tissue is found in sub-adults too (it would be more likely to be found in juveniles that are still growing), but it is not found in any growth stage of Paracyclotosaurus, which also lacks the woven-fibered bone of the smallest and most rapidly growing individuals of Cherninia. So Cherninia = fast grower, Paracyclotosaurus = slow grower. The histological / microanatomical differences can be correlated with distinct differences in the proportion of limb features and the torsion of the humerus and femur, suggesting biomechanical differences in their locomotion. The authors proposed that Cherninia was a classic obligately aquatic taxon, like what we think of most stereospondyls, whereas Paracyclotosaurus was more capable of moving around on land, not like what we typically think of large stereospondyls. It's debatable in my opinion whether the latter "spent a considerable amount of time on land," but it definitely seems likely that being able to at least move between ponds would have been advantageous, and it suggests that there may be a lot of cryptic information to be derived from the histology and microanatomy that is obscured by more conserved external anatomy.
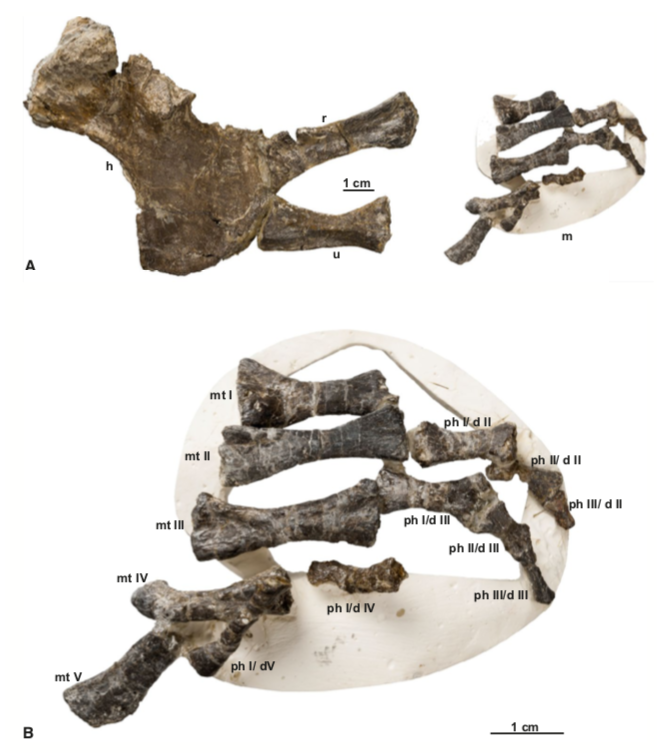
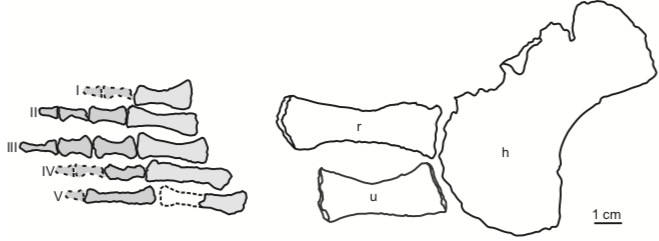
Comparison of sections of the humerus of Panthasaurus (on the left) with the femur, ulna, and tibia (on the right; A-H = femur; I-J = ulna; K-M = tibia). 2020 was the first year that I didn't have any new metoposaurid papers published (there's stuff in the pipeline, don't worry), but the metoposaurid histology train rolls on. Teschner et al. (PeerJ) reported histology and microanatomy of the Indian metoposaurid, Panthasaurus. This taxon shows pretty normal features for obligately aquatic stereospondyls, like parallel-fibered bone with remodelling that's accompanied by lamellar bone tissue (these reflect slower rates of growth), a fairly poorly vascularized cortex, and distinct growth marks (but not LAGs). This study is really nice because it adds to the growing body of literature helping us to understand how closely related taxa (within the same family) might differ in ecology or response to local climate, things we might not be able to nuance out from the external anatomy alone. Years ago, there was a nice comparative study by Dorota Konietzko-Meier (who's on this paper) and Nicole Klein comparing the signals in Dutuitosaurus from Morocco and Metoposaurus from Poland, which showed that the Polish paleoenvironment was milder and led to slow-downs or short stagnations but not long cessations in growth. Panthasaurus shows a similar signal to Metoposaurus (growth zones and annuli but no LAGs), indicating the Indian climate was also not too harsh. The authors also sampled a bunch of different elements, which is a key step forward to expanding the utility of non-limbs for histological studies by providing a reference point that isn't a humerus or a femur; if you wonder why nobody has done any of this work on North American taxa, it's because there are very few limb bones that are basically off-limits to destructive sampling right now (a number of people other than me have looked around, and this is why I keep cutting intercentra instead). Finally, Uliakhin et al. (Paleontol J) reported what is probably the highest age estimation for any Paleozoic vertebrate and definitely the highest one for any temnospondyl - a whopping 57 years! This was reported from the late Permian dvinosaur Dvinosaurus campbelli from Russia; dvinosaurs are one of the few uncontroversially fully aquatic Paleozoic temnospondyls, and there remains fringe speculation that one of the stereospondyl clades might actually be tied to this group. We see a lot of the same features that typify stereospondyls (e.g., parallel-fibered bone, lots of secondary remodeling from the medullary cavity, calcified cartilage), which is, at minimum, a reflection of what happens when you're neotenic and end up permanently living in the water. 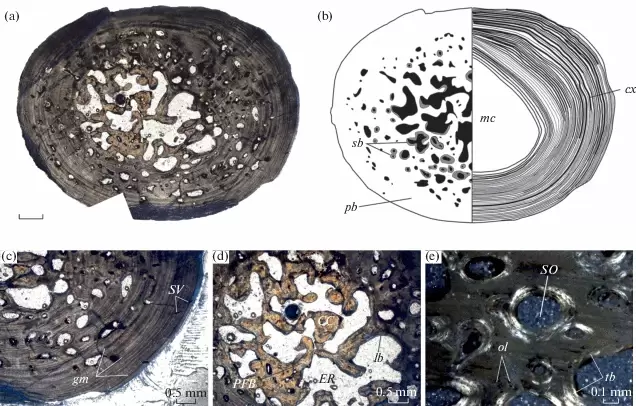 How about the 57 years - is that out of bounds? We know that slow-growing animals tend to live longer (why grow slow if you'll die in a year), and some modern amphibians top 60 years (and it's not one of those weird one-offs that far exceeds the norm for the species). Like the authors point out, neotenic individuals tend to live longer than metamorphosing ones. Of course, this assumes that the interpretation of the LAGs is correct; could it be double LAGs (two cessations in growth per year) for example (thus halving the estimated age)? Hard to say. The giveaway for double LAGs is their spacing - two closely spaced lines with a gap smaller than that from the line on either side. That doesn't mean you couldn't get double LAGs that are spatially indistinguishable from normal LAGs if the process that formed them was timed differently. For example, the way that we define seasons usually sets the start of winter and the start of summer as exactly 6 months apart. But as most people experience them, the "peak conditions" of each season aren't on Day 1, and the peaks may not be 6 months apart. Growing up in SoCal, I would say the hottest month of the year can be as late as September, and having lived in Canada, February or March can be the coldest. That asymmetry is likely what produces the characteristic spacing of double LAGs. But of course there's variation - who really knows the finer nuances of the climate in Russia over 250 million years ago, after all. So it's possible that climate peaks really could have been nearly 6 months apart and thus produce a pattern resembling single LAGs when the animal actually stopped growing twice a year. But we can't really prove that either. The best way to interpret this would be to cautiously accept the author's interpretation (single LAGs = 57 years) but not to make too much of it. Comparing maximum ages in extinct taxa is hard because how do you know that you got one that died around its maximum age? A lot of animals (especially those lower on the food chain) frequently die well before their maximum age, often because they're eaten, so the fact that other temnospondyls rarely exceed estimates of 15 years only says something about the sample, not necessarily the species as a whole. Piling up
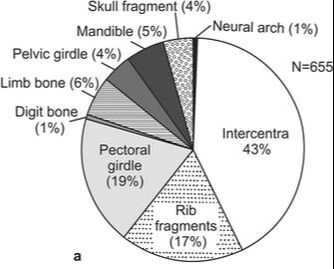
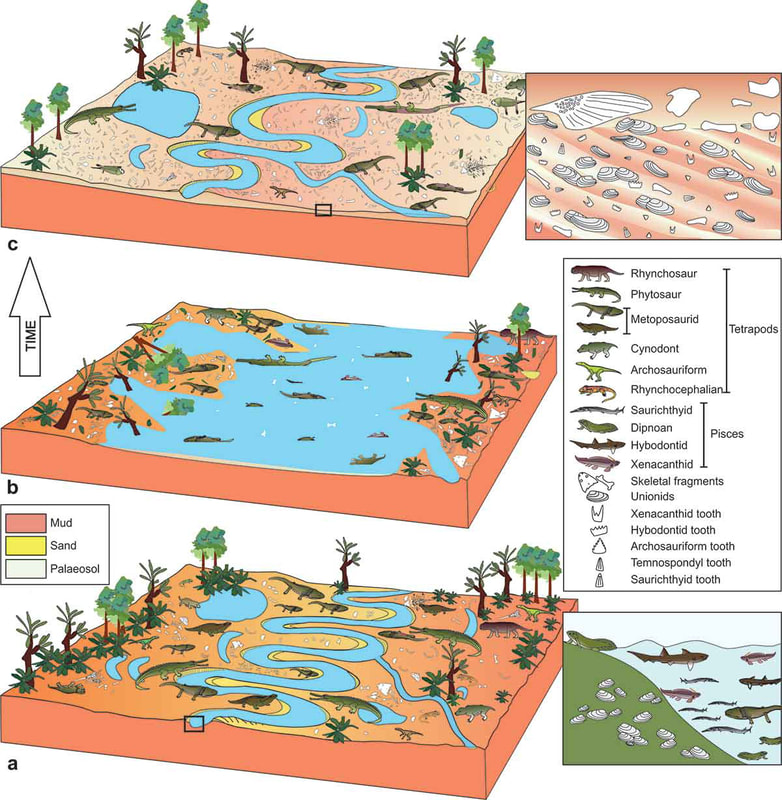
Based on a slew of taphonomic analyses, there appears to be disparate pre-burial conditions between the different tetrapods found at the site that explains the differences in their representation (the complex graph on the top left). There are mixed age classes for the metoposaurids, whereas the phytosaurs and rhynchosaurs are skewed towards either juveniles or adults. Obviously some of this has to do with which animals lived in the immediate environment versus which might have been carried in and the cause of death. This is not a mass death assemblage like that of Dutuitosaurus or the Lamy quarry insofar as it appears to be time-averaged for metoposaurids (but not rhynchosaurs, which may have mostly gotten swept in by a single event), and instead seems to be accumulation of their remains through periodic flooding and a build-up of remains in low-energy spots; the model is on the top right. The authors went so far as to argue that the metoposaurid remains might have been left out on the banks, exposed for years and constantly weathering, which might account for the total lack of complete skulls. Really great prospects based on the sheer volume of material that's come out of the site - hope to see more in the future! That's gotta hurtWith advances in technology and application of modern methods to paleontological specimens, paleopathology (studying diseases in fossils) has come a long way - we're seeing more and more reports of specific diagnoses and maladies beyond "that is clearly not right." Novikov et al. (Paleontol J) describe a bone lesion in the Early Triassic trematosaur Benthosuchus. This is one of the most common Early Triassic tetrapods found around the world (but mostly in Russia in this case), and the authors report a bone lesion on the lower jaw, which would be immediately identifiable to even non-scientists as a very odd-looking round protrusion from the side. 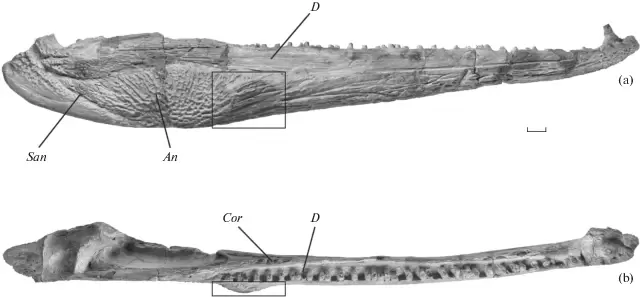 Based on features such as denser structure compared to surrounding bone, localized presence of a smooth feature, and no apparent connection to the teeth, the authors propose that this represents a non-odontogenic osteoma (non-tooth-related tumour). Because they couldn't do histology on the jaw, their results are based only on the external examination and the CT data (not the sharpest), so there are a few other possibilities (like a cyst or specifically a bone cancer). This remains the oldest example of a tumour forming in a tetrapod, although other reports like my friend Yara Haridy's 240-million-year-old turtle tumour are not that much later. Welcome to the club
A few other temnospondyl adjacent studies:
And the recent special issue on Karoo biozonation: https://pubs.geoscienceworld.org/sajg/issue/123/2 (note there are a few included taxa that are probably junior synonyms in some articles). In closingAll things considered, an excellent year for temnospondyl research! I reviewed a bunch of these papers as well, which is always neat to see what people are working on ahead of press. It's been a few months since I had anything come out (a coincidental pile-up of my 2019 productivity in the first half of this year), but I've got a few things working their way through the pipeline and will hopefully be getting back to some semblance of regular blogging in the new year! Thanks for reading, and best wishes for the new year! References
|
About the blogA blog on all things temnospondyl written by someone who spends too much time thinking about them. Covers all aspects of temnospondyl paleobiology and ongoing research (not just mine). Categories
All
Archives
January 2024
|
























 RSS Feed
RSS Feed
